Ketogenic amino acids are amino acids whose carbon skeleton can be fully or partially catabolized into acetoacetyl-CoA or acetyl-CoA, which serve as precursors for the synthesis of ketone bodies, hence the name, or for fatty acid synthesis.[1]
Amino acids are the fundamental constituents of proteins. Once their available pool is sufficient to meet the demands of protein synthesis, since the body does not have specific reserves of amino acids comparable to those of glycogen for glucose or triglycerides for fatty acids, it converts the excess into intermediates of the citric acid cycle, using them in the production of energy.[2]
When cellular energy requirements have also been met, the carbon residues resulting from amino acid catabolism are redirected towards the synthesis of glucose, fatty acids, or ketone bodies.[3]
Proteinogenic amino acids can be classified as ketogenic amino acids or glucogenic amino acids based on the metabolic fate of their carbon skeleton.[4] The carbon skeleton of ketogenic amino acids is catabolized to acetoacetyl-CoA and/or acetyl-CoA, while that of glucogenic amino acids is catabolized into one or more of the following five glucogenic precursor metabolites: pyruvate, oxaloacetate, α-ketoglutarate, succinyl-CoA and fumarate.[5]
However, this classification method is not unique as five amino acids are both ketogenic and glucogenic, since their catabolism originates at least one glucogenic precursor and one ketogenic precursor.[6]
Contents
- What are ketogenic amino acids?
- Biochemical basis
- Ketogenic amino acids during prolonged fasting
- Glyoxylate cycle
- References
What are ketogenic amino acids?
Only two of the twenty amino acids that make up proteins are exclusively ketogenic: leucine and lysine. The catabolism of their carbon skeleton results in the production of acetoacetyl-CoA and acetyl-CoA.[6]
| Aminoacid | Catabolite | ||
| ketogenic | glucogenic | ||
| Phenylalanine | Acetoacetyl-CoA | Fumarate | |
| Isoleucine | Acetyl-CoA | Succinyl-CoA | |
| Leucine * | Acetyl-CoA | Acetoacetyl-CoA | |
| Lysine * | Acetoacetyl-CoA | ||
| Tyrosine | Acetoacetyl-CoA | Fumarate | |
| Threonine | Acetyl-CoA | Succinyl-CoA | |
| Tryptophan | Acetyl-CoA | Acetoacetyl-CoA | Pyruvate |
| * Exclusively ketogenic amino acids | |||
Another five amino acids, namely isoleucine, phenylalanine, threonine, tryptophan, and tyrosine, are both ketogenic and glucogenic as their carbon skeleton catabolism leads to the formation of acetyl-CoA and/or acetoacetyl-CoA and a glucogenic precursor.[5]
The utilization of amino acid carbon skeletons is preceded by the removal of the amino group. Alanine and glutamate, glucogenic amino acids, play a key role in the transport of amino groups from extrahepatic tissues to the liver. In particular, alanine is transported from muscle and other peripheral tissues to the liver via the glucose-alanine cycle.[7]
Biochemical basis
Acetyl-CoA and acetoacetyl-CoA are not glucogenic precursors. The explanation lies in the stoichiometry of the citric acid cycle and the inability of animals to convert acetyl-CoA into pyruvate.[8]
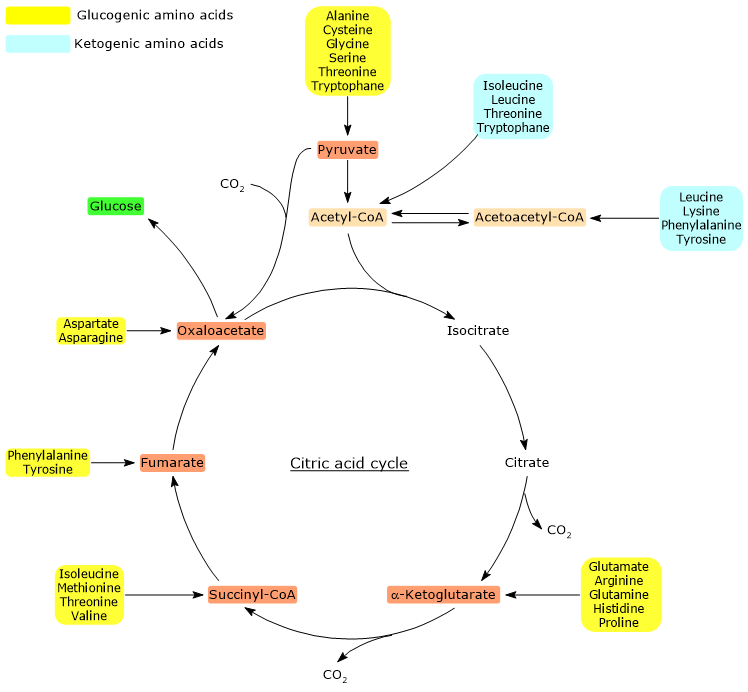 Acetyl-CoA enters the citric acid cycle through the reaction catalyzed by citrate synthase (EC 2.3.3.1). The enzyme catalyzes the condensation of acetyl-CoA with oxaloacetate to form citrate. Thus, a four-carbon compound is converted into a six-carbon compound, with a net gain of two carbon atoms.
Acetyl-CoA enters the citric acid cycle through the reaction catalyzed by citrate synthase (EC 2.3.3.1). The enzyme catalyzes the condensation of acetyl-CoA with oxaloacetate to form citrate. Thus, a four-carbon compound is converted into a six-carbon compound, with a net gain of two carbon atoms.
However, in the two subsequent oxidative decarboxylations of the cycle, catalyzed respectively by isocitrate dehydrogenase (EC 1.1.1.42) and α-ketoglutarate dehydrogenase multienzyme complex, two carbon atoms are lost.[4]
Therefore, the entry of acetyl-CoA into the cycle does not involve any net gain in carbon.[6]
The key factor is therefore the entry point of the carbon units, upstream or downstream of the oxidative decarboxylations of the citric acid cycle.[8]
Additionally, in animals, there is no metabolic pathway that allows the production of pyruvate from acetyl-CoA, due to the irreversibility of the reaction catalyzed by the pyruvate dehydrogenase complex, namely, the oxidative decarboxylation of pyruvate to acetyl-CoA.[8]
Ketogenic amino acids during prolonged fasting
Unlike glucogenic amino acids, ketogenic amino acids do not play a crucial role during prolonged fasting or in diets with severe carbohydrate restriction.
During prolonged fasting, the primary source of acetyl-CoA for ketogenesis is the β-oxidation of fatty acids, while the contribution of ketogenic amino acids is marginal. Under these conditions, glucogenic amino acids play a central role, as they ensure the maintenance of glucose homeostasis through gluconeogenesis when hepatic glycogen stores are depleted.[5]
Glyoxylate cycle
Plants, yeasts, and many bacteria can use acetyl-CoA for glucose synthesis because they have the glyoxylate cycle.
This cycle shares some reactions with the citric acid cycle but includes two unique reactions, catalyzed by isocitrate lyase (EC 4.1.3.1) and malate synthase (EC 2.3.3.9), and lacks decarboxylation reactions. Thus, organisms that possess the glyoxylate cycle can use fatty acids and ketogenic amino acids for glucose synthesis.[9]
References
- ^ D’Andrea G. Classifying amino acids as gluco(glyco)genic, ketogenic, or both. Biochem Educ 2000;28(1):27-28. doi:10.1016/s0307-4412(98)00271-4
- ^ Brosnan J.T. Interorgan amino acid transport and its regulation. J Nutr 2003;133(6 Suppl 1):2068S-2072S. doi:10.1093/jn/133.6.2068S
- ^ Litwack G. Human biochemistry. 2nd Edition. Academic Pr, 2021.
- ^ a b Moran L.A., Horton H.R., Scrimgeour K.G., Perry M.D. Principles of Biochemistry. 5th Edition. Pearson, 2012.
- ^ a b c Rosenthal M.D., Glew R.H. Medical Biochemistry – Human Metabolism in Health and Disease. John Wiley J. & Sons, Inc., 2009.
- ^ a b c Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 6th Edition. W.H. Freeman and Company, 2012.
- ^ Felig P., Pozefsk T., Marlis E., Cahill G.F. Alanine: key role in gluconeogenesis. Science 1970;167(3920):1003-1004. doi:10.1126/science.167.3920.1003
- ^ a b c Voet D. and Voet J.D. Biochemistry. 4th Edition. John Wiley J. & Sons, Inc. 2011.
- ^ Kondrashov F.A., Koonin E.V., Morgunov I.G., Finogenova T.V., Kondrashova M.N. Evolution of glyoxylate cycle enzymes in Metazoa: evidence of multiple horizontal transfer events and pseudogene formation. Biol Direct 2006;1:31. doi:10.1186/1745-6150-1-31