Nel caso in cui si consumi un eccesso di calorie in forma di carboidrati o proteine, tale surplus è utilizzato per la sintesi degli acidi grassi e quindi trigliceridi, cosa che non accade se l’eccesso di calorie proviene dai grassi stessi.
Indice
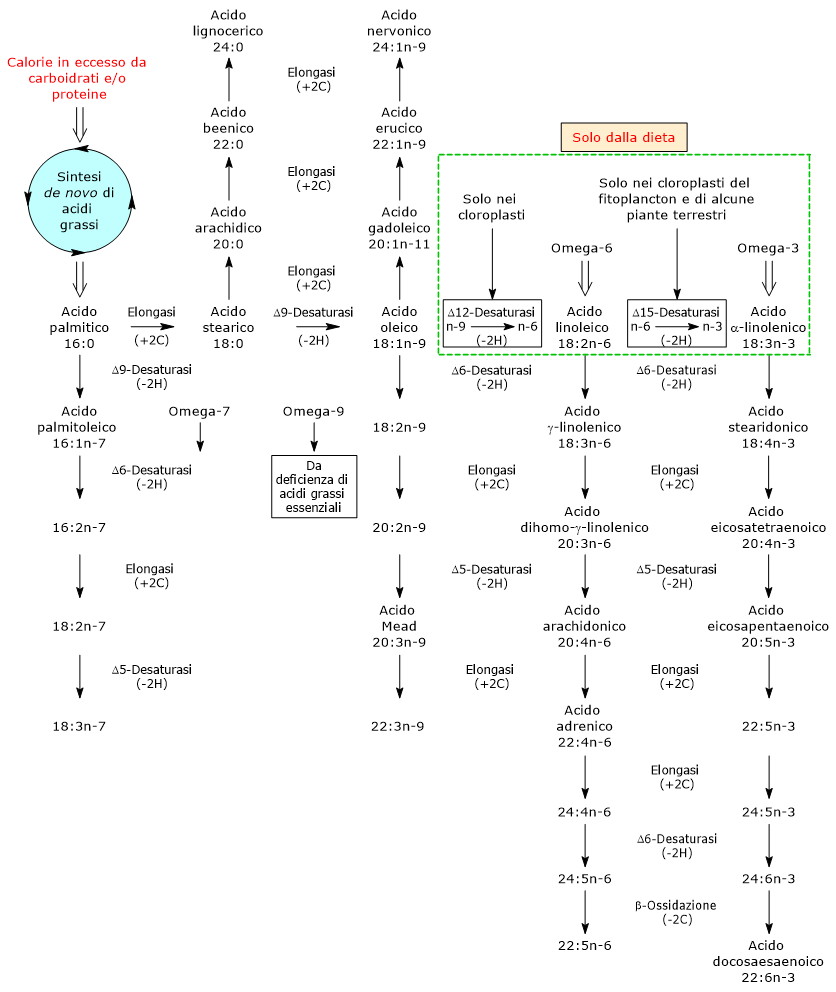
Sintesi de novo degli acidi grassi negli animali e piante
La sintesi de novo degli acidi grassi, molecole appartenenti al gruppo die lipidi, è in gran parte simile tra le piante e gli animali.
Avviene nei cloroplasti delle cellule fotosintetiche delle piante superiori, e nel citosol delle cellule animali, attraverso l’azione concertata di due enzimi: l’acetil-CoA carbossilasi (EC 6.4.1.2) e l’acido grasso sintetasi (EC 2.3.1.85).
L’acido grasso sintetasi catalizza una sequenza ripetuta di 4 passaggi tramite la quale la catena acilica è allungata di due atomi di carbonio, all’estremità carbossilica, a ogni passaggio attraverso il ciclo; questo processo in quattro tappe è il medesimo in tutti gli organismi.
Negli animali il sito primario per il metabolismo lipidico non è il tessuto adiposo ma il fegato. Il tessuto adiposo è comunque un organo importante in cui si verifica la sintesi degli acidi grassi, anche se negli esseri umani è meno attivo rispetto a molte altre specie animali.
Sebbene possano essere prodotti anche acido miristico, acido laurico e tracce di acido stearico, il prodotto principale della sintesi de novo sia nelle piante che negli animali è l’acido palmitico.
Da notare che in alcune piante, quali la palma e la noce di cocco, si verifica la terminazione della catena acilica prima che sia rilasciato l’acido palmitico: fino al 90% degli acidi grassi prodotti, e quindi presenti negli oli ricavati da questi vegetali, ha tra gli 8, atomi di carbonio, l’acido caprilico, un degli acidi grassi a catena media, e i 14 atomi di carbonio, ossia l’acido miristico.
Sintesi degli acidi grassi a catena lunga saturi e insaturi
L’acido palmitico è, tra gli acidi grassi saturi, il più comune nei lipidi di piante e animali, sebbene generalmente non sia presente in grandi quantità poiché può entrare in diverse vie metaboliche.
Infatti:
- è il precursore dell’acido stearico;
- può essere desaturato a dare acido palmitoleico, il precursore di tutti gli acidi grassi della famiglia omega-7, in una reazione catalizzata dall’enzima Δ9-desaturasi (EC 1.14.19.1). Questo enzima è ubiquitario sia nel regno animale che vegetale, è il più attivo del metabolismo lipidico nei tessuti di mammifero, ed è lo stesso che catalizza la desaturazione dell’acido stearico ad acido oleico.
La Δ9-desaturasi inserisce un doppio legame nella posizione 9-10 della catena carboniosa dell’acido grasso, posizione numerata a partire dall’estremità carbossilica della molecola. Se il substrato è l’acido palmitico, il doppio legame apparirà tra la posizione posizione n-7 e n-8 della catena, stavolta numerate a partire dall’estremità metilica, producendo acido palmitoleico, dunque il capostipite della famiglia omega-7.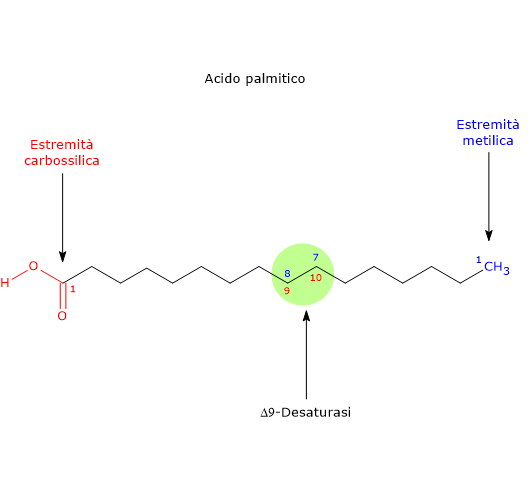 Se il substrato è l’acido stearico il doppio legame apparirà tra la posizione posizione n-9 e n-10 della catena, e verrà prodotto l’acido oleico.
Se il substrato è l’acido stearico il doppio legame apparirà tra la posizione posizione n-9 e n-10 della catena, e verrà prodotto l’acido oleico.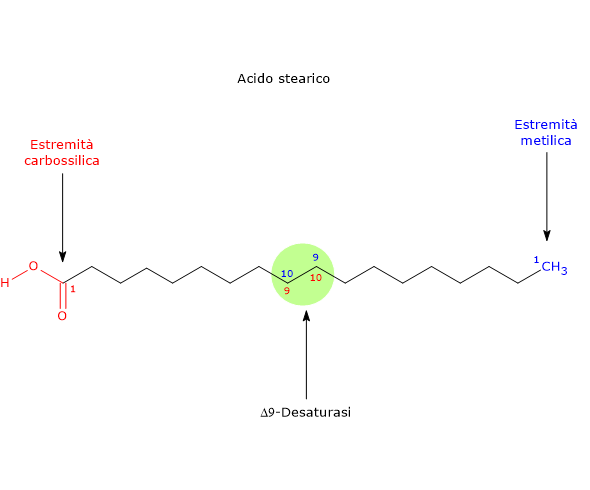
- Può essere esterificato in lipidi complessi.
Naturalmente nelle piante e negli animali ci sono acidi grassi sia con catene più lunghe sia con gradi di insaturazione maggiori rispetto a quelli visti, il tutto grazie a sistemi enzimatici (di nuovo di desaturazione e allungamento), che catalizzano reazioni di sintesi degli acidi grassi che sono organismo-, tessuto- e cellula specifiche.
Ad esempio l’acido stearico può essere:
- allungato ad acido arachidico, acido beenico e acido lignocerico, tutti acidi grassi saturi, in reazioni catalizzate da elongasi.
Anche in questo caso, l’allungamento della catena si verifica, sia nei mitocondri che nel reticolo endoplasmatico liscio, attraverso l’aggiunta di unità a due atomi di carbonio alla volta all’estremità carbossilica dell’acido grasso, tramite l’azione dei sistemi di allungamento degli acidi grassi (in particolare gli acidi grassi a catena lunga e molto lunga, da 18 a 24 atomi di carbonio, sono sintetizzati sulla superficie citosolica del reticolo endoplasmatico liscio); - desaturato, come visto, nella reazione catalizzata dalla Δ9-desaturasi, a dare acido oleico, un acido grasso omega-9.
Diversi ricercatori hanno supposto che la ragione per cui l’acido stearico non concorra allo sviluppo della ipercolesterolemia sia la sua rapida conversione in acido oleico.
L’acido oleico è il punto di partenza per la sintesi di molti acidi grassi insaturi attraverso reazioni di allungamento e/o desaturazione.
Infatti:
- l’allungamento della sua catena acilica per mezzo di un altro sistema enzimatico, l’acido grasso elongasi, produce acido gadoleico, acido erucico e acido nervonico, tutti acidi grassi monoinsaturi.
Gli acidi grassi saturi e i monoinsaturi della famiglia omega-9, in genere l’acido oleico, ma anche l’acido palmitoleico e gli altri omega-7, sono gli unici acidi grassi prodotti attraverso la sintesi de novo nei mammiferi. - Grazie all’azione consecutiva degli enzimi Δ12-desaturasi (1.14.19.6) e Δ15-desaturasi (EC 1.14.19.25), che inseriscono un doppio legame rispettivamente nella posizione posizione 12-13 e nella posizione 15-16 della catena carboniosa dell’acido grasso, l’acido oleico è convertito prima in acido linoleico, precursore di tutti gli acidi grassi polinsaturi omega-6, e quindi in acido alfa-linolenico, precursore di tutti gli acidi grassi polinsaturi omega-3 (acidi grassi polinsaturi che saranno prodotti a partire da questi precursori attraverso reazione ripetute di allungamento e desaturazione).
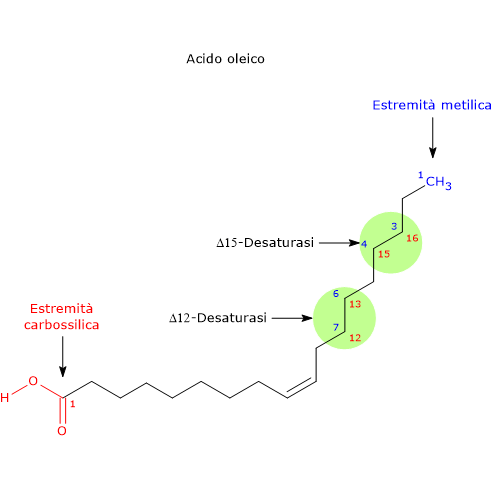
- In caso di deficienza di acidi grassi essenziali, l’acido oleico può essere desaturato e allungato a dare la serie di acidi grassi polinsaturi omega-9, con accumulo soprattutto di acido Mead.
Sintesi degli omega-3 e omega-6
I tessuti animali possono desaturare gli acidi grassi in posizione 9-10 della catena, grazie alla presenza della Δ9 desaturasi; come visto in precedenza se il substrato della reazione è l’acido palmitico il doppio legame apparirà tra le posizioni n-7 e n-8 della catena, con l’acido stearico tra le posizioni n-9 e n-10 portando così alla formazione rispettivamente di acido palmitoleico e oleico.
Gli animali non possiedono la Δ12- e la Δ15-desaturasi, enzimi in grado desaturare i legami carbonio carbonio oltre la posizione 9-10 della catena, e non sono quindi in grado di sintetizzare de novo gli acidi grassi polinsaturi omega-3 e omega-6, che posseggono doppi legami anche oltre la posizione 9-10, che sono pertanto acidi grassi essenziali.
La Δ12- e la Δ15-desaturasi sono presenti nelle piante; sebbene molte piante terrestri manchino della Δ15-desaturasi, detta anche omega-3 desaturasi, il plancton e le piante acquatiche delle acque fredde la posseggono e producono quantità abbondanti di acidi grassi omega-3.
Bibliografia
- Akoh C.C. and Min D.B. “Food lipids: chemistry, nutrition, and biotechnology”. CRC Press Taylor & Francis Group, 2008 3th ed. 2008
- Bender D.A. “Benders’ dictionary of nutrition and food technology”. 2006, 8th Edition. Woodhead Publishing. Oxford
- Burr G.O. and Burr M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. Nutr Rev 1973;31(8):148-149. doi:10.1111/j.1753-4887.1973.tb06008.x
- Chow Ching K. “Fatty acids in foods and their health implication”. 3rd Edition. CRC Press Taylor & Francis Group, 2008
- Rosenthal M.D., Glew R.H. Mediacal biochemistry. Human metabolism in health and disease. John Wiley & Sons, Inc. 2009
- Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3rd Edition. Elsevier health sciences, 2012