La corretta nutrizione è uno dei fattori chiave per il mantenimento di un buon stato di salute nel corso della vita.
Seguire una corretta alimentazione fin dalla nascita, con l’allattamento al seno, e in seguito durante l’infanzia, adolescenza e vita adulta permette di ottenere e mantenere un buon stato di salute. Quando associata a uno stile di vita salutare, concorre alla prevenzione dell’insorgenza di malattie croniche come le malattie cardiovascolari, l’osteoporosi, il diabete di tipo II, e molti tipi di tumori, condizioni sempre più diffuse nella moderna società.
Ed è assolutamente importante sottolineare la relazione tra nutrizione e microbiota intestinale, la comunità di microorganismi che colonizzano il nostro intestino. La dieta sembra infatti essere il fattore più importante nel determinarne la composizione, a partire dal primo alimento assunto, il latte materno.
La corretta nutrizione è l’intervento di elezione anche in presenza di reazioni allergiche a componenti degli alimenti, come nel caso della celiachia, condizione in cui l’unica cosa da fare è eliminare dall’alimentazione le fonti che apportano glutine per tutta la vita.
Nel caso di atleti, è essenziale, quando abbinata a un allenamento adeguato, per ottimizzare la performance in qualsiasi sport.
E tra i diversi tipi di diete, una delle più sane è la dieta mediterranea, un modello nutrizionale portato all’attenzione della comunità scientifica internazionale negli anni ’50 del secolo scorso dal lavoro di Ancel Keys, un fisiologo americano, in particolare con lo studio noto come il Seven Countries Study. La dieta mediterranea, ricca in prodotti vegetali, come l’olio extravergine di oliva, le verdure, i legumi, e i cereali integrali, e povera in carni rosse e prodotti derivati e latticini ad alto contenuto di grassi, assicura un ottimo apporto di composti antiossidanti e con azioni antiinfiammatorie, di fibre, e un basso apporto di grassi saturi.
L’ipertensione arteriosa viene definita come una pressione arteriosa media a riposo di 140/90 o maggiore e/o l’uso corrente di medicinali antiipertensivi.
E’ il problema di salute pubblica più comune nei paesi industrializzati. Spesso definita “killer silenzioso”, in quanto le persone affette possono essere asintomatiche per molti anni e poi andare incontro a un attacco di cuore fatale, è uno tra i maggiori fattori di rischio per lo sviluppo di malattia coronarica, infarto del miocardio, insufficienza cardiaca, ictus, e una delle principali cause di morbilità e mortalità. Tuttavia, tra i fattori di rischio di malattie cardiovascolari, è quello maggiormente modificabile.
L’ipertensione può essere primaria o essenziale e secondaria.
L’ipertensione primaria, responsabile di circa il 95% dei casi, è probabilmente conseguenza di fattori ambientali, fattori genetici, e della loro interazione. Tra i fattori ambientali la dieta ha un ruolo centrale. Tra i fattori genetici l’interesse si è focalizzato sui fattori che influenzano la risposta pressoria all’assunzione di sale, e sono stati identificati diversi genotipi, molti dei quali influenzano l’asse renina-angiotensina-aldosterone o il trattamento del sale a livello renale.
L’ipertensione secondaria è causata da altre patologie, solitamente endocrine, ad esempio l’ipertiroidismo, l’iperaldosteronismo, e la sindrome di Cushing.
Una pressione arteriosa al di sopra dei valori ottimali, anche se non ancora nell’intervallo ipertensivo o pre-ipertensivo, conferisce un aumento del rischio di malattie cardiovascolari, come mostrato dal fatto che quasi un terzo delle morti da malattia coronarica correlate alla pressione sono stimate accadere in soggetti non ipertesi, con pressione sistolica di 120-139 mm Hg o diastolica di 80-89 mm Hg. Ciò significa che il rischio di malattie cardiovascolari aumenta per tutto l’intervallo pressorio a partire da 115/75 mm Hg.
Categoria
Pressione arteriosa (mm Hg)
Sistolica
Diastolica
Ottimale
< 120
< 80
Normale
< 130
< 85
Normale – Alta
130 – 139
85 – 89
Ipertensione di grado 1
140 – 159
90 – 99
Ipertensione di grado 2
160 – 179
100 – 109
Ipertensione di grado 3
≥180
≥110
Ipertensione sistolica isolata
≥140
≤ 90
Infine, i soggetti pre-ipertesi hanno un elevato rischio, pari a circa il 90%, di sviluppare nel tempo l’ipertensione, sebbene il passaggio non sia inevitabile.
Ipertensione nella terza età
La prevalenza dell’ipertensione aumenta con l’aumentare dell’età, come mostrato dal fatto che più della metà della popolazione adulta oltre i 60 anni è ipertesa.
Il rischio correlato all’età è funzione di variabili legate allo stile di vita quali l’aumento di peso, la scarsa attività fisica, l’uso eccessivo di sale, grassi e acidi grassi saturi, alcool e ipercolesterolemia, e un basso apporto di frutta e verdura, piuttosto che all’invecchiamento in se. Ad esempio, studi condotti su vegetariani che vivono in paesi industrializzati hanno mostrato che tali abitudini alimentari sono associate con un minor aumento della pressione con l’aumentare dell’età, e con una pressione marcatamente più bassa se comparata a quella dei non vegetariani.
Ipertensione ed età pediatrica
Secondo uno studio condotto da un team di ricercatori della Johns Hopkins University la prevenzione dell’ipertensione ha inizio nell’infanzia.
Inoltre, una meta-analisi su studi condotti su diverse popolazioni, pubblicati tra gennaio 1970 e luglio 2006, ha esaminato il cammino della pressione dall’infanzia all’età adulta dimostrando che la pressione dell’infanzia è associata con quella nei periodi successivi della vita, e che un valore elevato nell’infanzia è probabilmente di aiuto nel predire l’ipertensione in età avanzata.
Infine, altri studi hanno anche dimostrano che l’aumentata pressione arteriosa tra i bambini è correlata alla crescente epidemia di obesità.
Come prevenire l’ipertensione
Nelle ultime due decadi negli USA è stato documentato un trend discendente della pressione arteriosa, cui ha contribuito l’adozione di uno stile di vita più salutare.
Le modificazioni dello stile di vita che riducono efficacemente la pressione sono:
limitare l’apporto di sale e di altre forme di sodio;
seguire un’alimentazione ricca di frutta fresca, verdura, carboidrati complessi e prodotti caseari a basso contenuto di grassi;
aumentare l’apporto di potassio consumando frutta, verdura e legumi;
perdere peso corporeo se in sovrappeso, o prevenirne l’aumento tra coloro che sono magri;
aumentare l’attività fisica di bassa o moderata intensità;
smettere di fumare;
Questi cambiamenti rappresentano la prima linea di difesa nella lotta contro la pressione alta, e non dovrebbero essere fatti uno alla volta: i migliori risultati si ottengono quando i cambiamenti avvengono contemporaneamente, come dimostrato da due studi in cui interventi “multicomponent” hanno ridotto la pressione arteriosa nei partecipanti ipertesi e non ipertesi.
Infine, è stato dimostrato una relazione anche tra assunzione di alcool e ipertensione.
Ruolo del potassio
Il potassio, un nutriente essenziale per l’uomo, è il più abbondante catione presente nei fluidi intracellulari. E’ quindi ampiamente diffuso nei cibi che derivino da tessuti animali e vegetali, ma che non abbiano subito processi di salatura e/o essicazione. Anche i metodi di cottura tendono a ridurne la quantità. Considerando le verdure, il peggior metodo di cottura è la bollitura in abbondante acqua, prolungata per più di un’ora, mentre il migliore è la cottura a microonde.
Potassio
Contenuto
>250 mg/100 g
Legumi
Legumi secchi, quali ceci, fagioli, lenticchie, piselli e soia, e fagioli freschi.
Carne/prodotti della pesca, sia freschi che conservati. Questi ultimi sono da limitare a causa dell’elevato contenuto in sodio.
Un’elevata assunzione di potassiocon gli alimenti e la pressione arteriosa sono inversamente correlate, come dimostrato da studi condotti su animali, studi epidemiologici osservazionali, sperimentazioni cliniche, studi di alimentazione controllata, come il DASH Study e l’OmniHeart trial, e meta-analisi. Inoltre, un elevato apporto di potassio aumenta anche l’escrezione urinaria di sodio.
La strategia ottimale per aumentare l’apporto di potassio è quella di consumare alimenti naturalmente ricchi del minerale quali frutta e verdura di stagione, e legumi, alimenti tipici della dieta mediterranea. Non è quindi difficile raggiungere il livello di assunzione giornaliera raccomandata, per la popolazione sana, che è pari a 4,7 g/die.
Ruolo del sodio
A differenza del potassio, il sodio è il principale catione dei liquidi extracellulari, di cui ne condiziona fortemente il valore della pressione osmotica.
Le principali fonti alimentari sono tre.
La più intuitiva è il sale utilizzato a tavola, che rappresenta fino al 20% dell’apporto giornaliero. Si noti che, sebbene molto spesso i termini sale e sodio siano usati in modo interscambiabile, ciò non è corretto. Sulla base del peso, il sale è costituito per il 40% da sodio e per il restante 60% da cloro.
Una seconda fonte è il sale o i composti del sodio aggiunti durante la preparazione o la trasformazione dei cibi. Tra il 35 e 80% del sodio assunto giornalmente proviene dai cibi trasformati, quali:
carne e pesce affumicati, sott’olio o sotto sale;
estratti di carne e salse, snack salati, la salsa di soia, e la salsa piccante;
Trascurabile è invece la terza fonte, il sodio naturalmente presente negli alimenti, in genere basso negli alimenti freschi.
Un elevata assunzione di sodio contribuisce all’aumento della pressione arteriosa e allo sviluppo dell’ipertensione. Ciò è supportato da molti studi epidemiologici, su animali, di migrazione delle popolazioni, e meta-analisi, con la prova finale derivante da studi accuratamente controllati dose-risposta. Inoltre, nelle società primitive, dove l’apporto di sodio è molto basso, le persone raramente vanno incontro a ipertensione, e con l’avanzare dell’età non si verifica l’aumento della pressione arteriosa.
Pertanto, per prevenire lo sviluppo dell’ipertensione si raccomanda una riduzione del suo apporto. In ragione degli alimenti a disposizione e dell’elevato consumo corrente, una raccomandazione ragionevole può essere di porre come limite superiore un apporto di sodio di 2,3 g/die, pari a 5,8 g/die di sale.
Come può essere raggiunto questo valore?
Usando meno sale possibile durante la preparazione dei cibi.
Evitando di aggiungere sale una volta che i cibi sono in tavola.
Limitando il consumo di cibi trasformati molto salati.
Studi clinici hanno documentato che una ridotta assunzione di sodio è in grado di abbassare la pressione arteriosa anche in un quadro di assunzione di farmaci antipertensivi, e può facilitare il controllo dell’ipertensione stessa.
Alcuni componenti della dieta possono modificare la risposta pressoria al sodio. Un elevato apporto dietetico di cibi ricchi di potassio e calcio può prevenire o attenuare l’aumento della pressione a seguito di un dato aumento dell’apporto di sodio. Al contrario, alcuni dati, osservati principalmente in modelli animali, suggeriscono che una elevata assunzione di saccarosio potrebbe potenziare la sensibilità della pressione arteriosa verso sodio.
Nota: elevati apporti di sodio possono contribuire allo sviluppo dell’osteoporosi, in quanto determinano un aumento dell’escrezione renale di calcio, in modo particolare se l’assunzione giornaliera di calcio è bassa.
Ruolo del peso corporeo
Il peso corporeo, in particolare il sovrappeso e l’obesità, sono un fattore determinante della pressione arteriosa a ogni età. Infatti:
è stato stimato che il rischio di sviluppare una pressione elevata è da due a sei volte maggiore nelle persone in sovrappeso rispetto ai normopeso;
c’è una correlazione lineare tra la pressione arteriosa e il peso corporeo o l’indice di massa corporea, che, se maggiore di 27, è correlato con un aumento della pressione;
anche quando l’assunzione di sodio è mantenuta costante, la correlazione tra variazione di peso e pressione arteriosa è lineare;
il 60% dei soggetti affetti da ipertensione è più del 20% in sovrappeso;
la distribuzione centrale della massa grassa, come fattore determinante l’aumento della pressione arteriosa, con una circonferenza addominale superiore a 88 cm nelle donne e 102 negli uomini, è più importante della localizzazione periferica del grasso stesso, sia negli uomini che nelle donne;
la perdita di peso, sia nei soggetti affetti da ipertensione che nei normotesi, può ridurre la pressione arteriosa, e la riduzione si verifica prima, e senza, il raggiungimento di un peso corporeo desiderabile.
Ruolo dell’attività fisica
L’attività fisica produce un calo della pressione sistolica e diastolica. Quindi, per la prevenzione primaria dell’ipertensione è importante aumentare l’attività fisica di bassa o moderata intensità per 30-45 minuti per 3-4 volte alla settimana fino a un’ora quasi tutti i giorni, come raccomandato dalla Organizzazione Mondiale della Sanità. Di contro, le persone meno attive hanno dal 30 al 50% di probabilità in più di sviluppare ipertensione rispetto a quelle attive.
Bibliografia
Appel L.J., Brands M.W., Daniels S.R., Karanja N., Elmer P.J. and Sacks F.M. Dietary approaches to prevent and treat HTN: a scientific statement from the American Heart Association. Hypertension 2006;47:296-08. doi:10.1161/01.HYP.0000202568.01167.B6
Bibbins-Domingo K., Chertow G.M., Coxson P.G., Moran A., Lightwood J.M., Pletcher M.J., and Goldman L. Projected effect of dietary salt reductions on future cardiovascular disease. N Engl J Med 2010;362:590-9. doi:10.1056/NEJMoa0907355
Cappuccio FP. Overview and evaluation of national policies, dietary recommendtions and programmes around the world aiming at reducing salt intake in the population. World Health Organization. Reducing salt intake in populations: report of a WHO forum and technical meeting. WHO Geneva 2007;1-60.
Chen J, Gu D., Jaquish C.E., Chen C., Rao D.C., Liu D., Hixson J.E., Lee Hamm L., Gu C.C., Whelton P.K. and He J. for the GenSalt Collaborative Research Group. Association between blood pressure responses to the cold pressor test and dietary sodium intervention in a chinese population. Arch Intern Med. 2008;168:1740-1746. doi:10.1001/archinte.168.16.1740
Chen X. and Wang Y. Tracking of blood pressure from childhood to adulthood. A systematic review and meta-regression analysis. Circulation 2008;117:3171-80. doi:10.1161/CIRCULATIONAHA.107.730366
Denton D., Weisinger R., Mundy N.I., Wickings E.J., Dixson A., Moisson P., Pingard A.M., Shade R., Carey D., Ardaillou R., Paillard F., Chapman J., Thillet J. & Michel J.B. The effect of increased salt intake on blood pressure of chimpanzees. Nature Med 1995;10:1009-1016. doi:10.1038/nm1095-1009
Ford E.S., Ajani U.A., Croft J.B., Critchley J.A., Labarthe D.R., Kottke T.E., Giles W.H, and Capewell S. Explaining the decrease in U.S. deaths from coronary disease, 1980-2000. N Engl J Med 2007;356:2388-98. doi:10.1056/NEJMsa053935
Geleijnse J.M., Witteman J.C., den Breeijen J.H., Hofman A., de Jong P., Pols H.A. and Grobbee D.E. Dietary electrolyte intake and blood pressure in older subjects: the Rotterdam Study. J Hyperten 1996;14:73741. doi:10.1097/00004872-199606000-00009
Gutiérrez O.M. Sodium- and phosphorus-based food additives: persistent but surmountable hurdles in the management of nutrition in chronic kidney disease. Adv Chronic Kidney Dis 2013;20(2):150-6. doi:10.1053/j.ackd.2012.10.008
Harlan W.R. and Harlan L.C. Blood pressure and calcium and magnesium intake. In: Laragh J.H., Brenner B.M., eds. Hypertension: pathophysiology, diagnosis and management. 2end ed. New York: Raven Press 1995;1143-1154
He F.J., Tan M., Ma Y., MacGregor G.A. Salt reduction to prevent hypertension and cardiovascular disease: JACC state-of-the-art review. J Am Coll Cardiol 2020;75(6):632-647. doi:10.1016/j.jacc.2019.11.055
Holmes E., Loo R.L., Stamler J., Bictash M., Yap I.K.S., Chan Q., Ebbels T., De Iorio M., Brown I.J., Veselkov K.A., Daviglus M.L., Kesteloot H., Ueshima H., Zhao L., Nicholson J.K. and Elliott P. Human metabolic phenotype diversity and its association with diet and blood pressure. Nature 2008;453:396-400. doi:10.1038/nature06882
Nugroho P., Andrew H., Kohar K., Noor C.A., Sutranto A.L. Comparison between the world health organization (WHO) and international society of hypertension (ISH) guidelines for hypertension. Ann Med 2022;54(1):837-845. doi:10.1080/07853890.2022.2044510
Sesso H.D., Cook N.R., Buring J.E., Manson J.E. and Gaziano J.M. Alcohol consumption and the risk of hypertension in women and men. Hypertension 2008;51:1080-1087. doi:10.1161/HYPERTENSIONAHA.107.104968
Simpson F.O. Blood pressure and sodium intake. In: Laragh J.H., Brenner B.M. eds. Hypertension: pathophysiology, diagnosis and management. 2end ed. New York: Raven Press 1995;273-281
Stone M.S., Martyn L., Weaver C.M. Potassium intake, bioavailability, hypertension, and glucose control. Nutrients 2016;8(7):444. doi:10.3390/nu8070444
Strazzullo P., D’Elia L., Kandala N. and Cappuccio F.P. Salt intake, stroke, and cardiovascular disease: meta-analysis of prospective studies. BMJ 2009;339:b4567. doi:10.1136/bmj.b4567
Tzoulaki I., Brown I.J., Chan Q., Van Horn L., Ueshima H., Zhao L., Stamler J., Elliott P., for the International Collaborative Research Group on Macro-/Micronutrients and Blood Pressure. Relation of iron and red meat intake to blood pressure: cross sectional epidemiological study. BMJ 2008;337:a258. doi:10.1136/bmj.a258
Unger T., Borghi C., Charchar F., Khan N.A., Poulter N.R., Prabhakaran D., Ramirez A., Schlaich M., Stergiou G.S., Tomaszewski M., Wainford R.D., Williams B., Schutte A.E. 2020 International society of hypertension global hypertension practice guidelines. hypertension 2020;75(6):1334-1357. doi:10.1161/HYPERTENSIONAHA.120.15026
Weinberger M.H. The effects of sodium on blood pressure in humans. In: Laragh J.H., Brenner B.M., eds. Hypertension: pathophysiology, diagnosis and management. 2end ed. New York: Raven Press 1995;2703-2714.
Writing Group of the PREMIER Collaborative Research Group. Effects of comprehensive lifestyle modification on blood pressure control: main results of the PREMIER Clinical Trial. JAMA 2003;289:2083-2093. doi:10.1001/jama.289.16.2083
World Health Organization, International Society of Hypertension Writing Group. 2003 World Health Organization (WHO)/ISH statement on management of HTN. Guidelines and recommendations. J Hyperten 2003;21:1983-92. doi:10.1097/00004872-200311000-00002
E il colesterolo assunto con gli alimenti?
Non esiste una correlazione diretta tra i livelli di colesterolo plasmatico e l’assunzione di colesterolo con gli alimenti. Tuttavia, il colesterolo alimentare può aumentare i livelli del colesterolo plasmatico se assunto in associazione agli acidi grassi saturi e agli acidi grassi trans.
Se comunque vuoi ridurre l’assunzione di colesterolo è consigliabile diminuire l’uso di prodotti di origine animale e/o consumare latte parzialmente scremato o scremato, formaggi con basso contenuto di grassi, yogurt magro, e carne magra.
Un fattore di rischio per l’ipercolesterolemia è rappresentato da un’elevata assunzione di acidi grassi saturi, un gruppo di lipidi che possono essere facilmente utilizzati per la produzione endogena di colesterolo.
Questi acidi grassi sono presenti nella carne, nei latticini, e in elevata quantità nei grassi e oli vegetali, come la margarina, l’olio di palma, l’olio di semi di palma, e l’olio di cocco, molto utilizzati nell’industria dolciaria.
Cosa fare:
eliminare il grasso visibile dalla carne o acquistarne tagli magri;
sostituire il latte intero, il burro, i formaggi grassi, le creme, e i gelati con prodotti con un contenuto in grassi più basso, come lo yogurt magro, il latte parzialmente scremato o scremato, e i formaggi magri;
evitare i prodotti di pasticceria.
Acidi grassi trans
Gli acidi grassi trans o grassi trans costituiscono un fattore di rischio molto elevato, e non solo per l’ipercolesterolemia.
Studi condotti su volontari ne hanno evidenziato una grande capacità aterogenica dovuta a modificazione a carico delle lipoproteine plasmatiche, dove si osserva una diminuzione dei livelli delle HDL e un aumento dei livelli delle LDL e dei trigliceridi.
Dove si possono trovare?
In molti alimenti per bambini.
Nei prodotti da forno industriali come cracker, grissini, biscotti, pane confezionato, e snack.
In prodotti come zuppe, piatti pronti, freschi o surgelati, e in paste di base per torte e pizze casalinghe.
Nei dadi da brodo.
Nelle caramelle morbide.
In alcuni cereali per la prima colazione.
Nei gelati, nei surrogati vegetali della panna, e nella margarina.
In molte conserve vegetali, incluse le marmellate.
Per quanto riguarda il contenuto in acidi grassi saturi e trans, molto spesso non ci sono differenze tra i prodotti classici e quelli “naturali” o “biologici”.
Cosa fare?
Evitare di acquistare prodotti che nella composizione presentino grassi/oli vegetali idrogenati o parzialmente idrogenati, ed evitare di acquistare prodotti fritti.
Sovrappeso
Un significativo aumento della massa grassa contribuisce all’ipercolesterolemia.
Per molte persone la riduzione dell’assunzione degli acidi grassi saturi e trans non si accompagna a una riduzione della colesterolemia fino a che non c’è un concomitante calo del peso corporeo.
Cosa fare per tenere sotto controllo il peso?
Ridurre il consumo di grassi di origine animale e vegetale.
Ridurre i cibi ricchi di zuccheri semplici come dolci, soft drinks, dessert, caramelle e biscotti.
Non reintrodurre le calorie eliminate nei due punti precedenti con un eccessivo consumo di olio di oliva e amido, ossia pasta, patate, riso, pane.
Aumentare l’attività fisica.
Aumentare il consumo di frutta e verdura.
Cause genetiche
In questo caso è necessario un intervento farmacologico, che dovrà essere prescritto dal proprio medico curante, ma che deve essere comunque associato a una alimentazione adeguata.
Bibliografia
Ascherio A., Katan M.B., Zock P.L., Stampfer M.J., Willett W.C. Trans fatty acids and coronary heart disease. N Engl J Med 1999;340:1994-1998. doi:10.1056/NEJM199906243402511
Benito-Vicente A., Uribe K.B., Jebari S., Galicia-Garcia U., Ostolaza H., Martin C. Familial hypercholesterolemia: the most frequent cholesterol metabolism disorder caused disease. Int J Mol Sci 2018;19(11):3426. doi:10.3390/ijms19113426
Fernandez M.L., Murillo A.G. Is there a correlation between dietary and blood cholesterol? Evidence from epidemiological data and clinical interventions. Nutrients 2022;14(10):2168. doi:10.3390/nu14102168
Hu F.B., Willett W.C. Optimal diet for prevention of coronary heart disease JAMA 2002;288:2569-2578. doi:10.1001/jama.288.20.2569
Lichtenstein A.H. Dietary fat, carbohydrate, and protein: effects on plasma lipoprotein patterns J. Lipid Res. 2006;47:1661-1667. doi:10.1194/jlr.R600019-JLR200
Lichtenstein A.H., Ausman L., Jalbert S.M., Schaefer E.J. Effect of different forms of dietary hydrogenated fats on serum lipoprotein cholesterol levels. N Engl J Med 1999;340:1933-1940. doi:10.1056/NEJM199906243402501
Mensink R.P., Katan M.B. Effect of dietary trans fatty acids on high-density and low-density lipoprotein cholesterol levels in healthy subjects. N Engl J Med 1990;323:439-445. doi:10.1056/NEJM199008163230703
Mozaffarian D., Katan M.B., Ascherio A., Stampfer M.J., Willett W.C. Trans fatty acids and cardiovascular disease. N Engl J Med 2006;354:1601-1613. doi:10.1056/NEJMra054035
Shils M.E., Olson J.A., Shike M., Ross A.C.: “Modern nutrition in health and disease” 9th ed., by Lippincott, Williams & Wilkins, 1999
Il concetto di dieta mediterranea è stato introdotto negli anni ’50 dello scorso secolo dal fisiologo americano Ancel Benjamin Keys.
Keys per primo identificò la relazione tra dieta e insorgenza delle malattie cardiovascolari o CVD, in particolare grazie allo studio epidemiologico noto come Seven Countries Study, il primo studio a evidenziare questo modello alimentare.
La dieta mediterranea, che è riconosciuta come uno dei modelli alimentari più sani, è ricca di alimenti vegetali minimamente lavorati, quali verdure, legumi, cereali, meglio se integrali, con l’olio di oliva extravergine come principale fonte di lipidi. Quindi è un modello nutrizionale ricco in sostanze antiossidanti e con azione antiinfiammatoria.
Dopo lo Studio di Keys, molti altri gruppi di ricerca hanno evidenziato, in popolazioni appartenenti a paesi industrializzati e non, il ruolo protettivo di questo tipo di alimentazione non solo nei confronti della CVD, ma anche di malattie cronico-degenerative, dei disturbi depressivi, nonché una correlazione con miglioramenti sulla capacità di apprendimento. Ed è stato dimostrato che una maggiore aderenza alla dieta mediterranea è associata al miglioramento dello stato di salute e alla riduzione della mortalità in generale
Per tutto questo, non esiste associazione scientifica che sostenga la tesi secondo cui sia dannosa per la nostra salute.
In aggiunta, grazie al suo ridotto consumo di carni, è in grado di migliorare la salute pubblica anche concorrendo alla riduzione delle emissione dei gas serra.
In definitiva, la dieta mediterranea rappresenta un tipo di alimentazione che deve essere salvaguardata e promossa, in contrapposizione al trend mondiale verso una uniformità dietetica che sta avanzando anche nei paesi del bacino mediterraneo.
Keys individuò la correlazione tra dieta e il rischio di sviluppare malattie cardiovascolari nei primi anni ’50 del secolo scorso confrontando i tassi di insorgenza di CVD tra dirigenti aziendali americani e popolazioni europee appena uscite dalla seconda guerra mondiali. Mentre tra i primi, soggetti ben nutriti, i tassi di insorgenza erano elevati, nei secondi, che invece si trovavano in una fase di indigenza alimentare, i tassi di insorgenza di CVD erano bassi. Queste osservazioni portarono Keys a ipotizzare una correlazione tra il contenuto di grassi nella dieta e i decessi per malattie cardiovascolari.
La successiva osservazione di una frequenza estremamente bassa di malattie coronariche e certi tipi di tumore nella popolazione dell’isola di Creta, in molta della restante popolazione greca e in quella del sud dell’Italia rispetto a quanto osservato negli USA, portò Keys a ipotizzare che l’alimentazione di quelle popolazioni, caratterizzata da un basso contenuto di grassi di origine animale, costituisse un fattore di protezione.
Il passo successivo fu l’avvio di quello che sarà conosciuto come il Seven Countries Study, uno studio epidemiologico osservazionale a lungo termine, ben 25 anni, e lo studio più conosciuto sulla dieta mediterranea, nel quale fu osservata una correlazione inversa tra la dieta e il rischio di morte generale e legata a malattie cardiovascolari. Lo Studio mostrò che i grassi saturi sono il principale fattore di rischio dietetico e che seguire un’alimentazione di tipo mediterraneo portava a un riduzione del rischio di insorgenza di malattie cardiovascolari.
Caratteristiche della dieta mediterranea
La dieta mediterranea è un modello nutrizionale caratterizzato dal consumo prevalente di cibi di origine vegetale quali legumi, verdure, cereali integrali e olio extravergine d’oliva, il che assicura un buon apporto di fibre, antiossidanti, fitosteroli, polifenoli e acidi grassi insaturi.
Per quanto riguarda i prodotti di origine animale, il consumo di carne, in particolare carne rossa e prodotti derivati, nonché di prodotti caseari ad alto contenuto di grassi dovrebbe essere limitato, mentre dovrebbero essere presenti pesci e frutti di mare.
Il consumo di etanolo dovrebbe essere moderato, da assumersi con il vino rosso durante i pasti.
E tra i greci che hanno partecipato allo studio EPIC, l’olio extravergine di oliva, le verdure, i legumi, un modesto apporto di etanolo, assieme a un basso consumo di carne e derivati sono risultati predittori di una minore mortalità.
Pietra angolare della dieta mediterranea è l’olio extravergine di oliva. Da utilizzarsi preferibilmente crudo, è un’ottima fonte di acidi grassi monoinsaturi e contiene oltre 2000 diverse molecole, molte con attività antiossidante.
Rappresentando la principale fonte di lipidi, quando associato a un basso consumo di prodotti di origine animale ad alto contenuto di grassi, assicura un elevato rapporto tra acidi grassi monoinsaturi e acidi grassi saturi, il che concorre al miglioramento del profilo lipidico e del controllo glicemico nei diabetici.
E’ tuttavia fuorviante focalizzare l’attenzione su un singolo componente della dieta mediterranea; non esiste il “proiettile magico”, come mostrato da studi che si sono concentrati su un unico nutriente. Le persone non mangiano un singolo e isolato nutriente, ma un complesso di nutrienti e, cosa ancora più importante, i nutrienti interagiscono tra di loro in modo sinergistico o antagonistico. Quindi, i benefici per la salute apportati dalla dieta mediterranea sono dovuti a tutti i suoi componenti.
Dieta mediterranea e malattie croniche
Dopo il Seven Countries Study numerosissimi studi hanno dimostrato l’efficacia di questo modello nutrizionale nella prevenzione sia primaria che secondaria delle principali malattie croniche, dalle malattie cardiovascolari ai disturbi depressivi, nonché una riduzione della mortalità generale.
Di seguito alcuni esempi tra i moltissimi disponibili tra le pubblicazioni scientifiche.
Una meta-analisi ha analizzato l’associazione tra l’aderenza alla dieta mediterranea, mortalità, e incidenza di malattie dimostrando che “la maggiore aderenza alla dieta mediterranea è associata in modo significativo con una riduzione della mortalità generale, mortalità cardiovascolare, incidenza e mortalità per cancro, e incidenza del morbo di Parkinson.”
Uno studio multicentrico randomizzato ha dimostrato l’efficacia nella prevenzione primaria degli eventi cardiovascolari in soggetti ad alto rischio cardiovascolare.
E’ correlata sia a un minor rischio di morbo di Alzheimer che con il suo decorso e i suoi esiti: la maggior aderenza è associata con una minore mortalità e si suggerisce un effetto dose-risposta.
Ci sono prove crescenti che indicano un effetto protettivo sull’aumento di peso.
E’ stata riporta un’associazione inversa tra l’aderenza a queste abitudini dietetiche e l’incidenza del diabete di tipo 2 tra soggetti inizialmente in salute e in pazienti sopravvissuti a un infarto miocardico.
E’ associata a una minor prevalenza della sindrome metabolica.
Studi epidemiologici e di intervento hanno rivelato un effetto protettivo nei confronti dell’infiammazione cronica lieve e delle sue complicanze metaboliche.
Ci sono prove che l’aderenza al modello della dieta mediterranea possa avere un potenziale ruolo protettivo per quanto riguarda la prevenzione dei disturbi depressivi.
Ruolo nella riduzione dei gas serra
La dieta mediterranea è in grado di migliorare la salute pubblica anche concorrendo alla riduzione dell’emissione dei gas serra, ossia anidride carbonica o CO2, metano, ossido di azoto e simili, provenienti dal settore zootecnico, responsabile dei 4/5 delle emissioni legate all’agricoltura. Queste emissioni sono maggiori di quelle dovute ai trasporti, e seconde solo a quelle derivanti dalla produzione di energia. Se a questo si aggiunge che la popolazione mondiale sta crescendo, e tale crescita è accompagnata da un aumento del consumo pro capite di carne, con stime secondo cui entro il 2030 si avrà un incremento della produzione di carne dell’85% rispetto al 2000, il ruolo della dieta mediterranea nei confronti della riduzione dell’emissione dei gas serra diviene ancora più evidente.
Allevamento bovino e gas serra
Analizzando nel dettaglio i gas serra derivanti dall’allevamento bovino, il maggiore responsabile dell’emissione nel comparto zootecnico, questi derivano come di seguito indicato:
40% dalla perdita di piante annuali, erbe e alberi che ricoprirebbero il terreno dove si coltiva il foraggio;
32% dalle emissioni di metano da parte dei rifiuti animali e dagli animali stessi;
14% dai fertilizzanti per la coltivazione dei cereali da foraggio, di cui ne sono necessari 16 chili per chilo di carne consumata;
14% dalla produzione agricola generale.
Nella tabella sono confrontate le emissioni di CO2 dovute alla produzione di diversi cibi, considerando porzioni da 225 g, con quelle rilasciate da una macchina a benzina che fa circa 12 chilometri con un litro.
Alimenti
Distanza
Grammi di CO2 equivalente
Patate
300 metri
59
Mele
320 metri
68
Asparagi
440 metri
91
Pollo
1,17 chilometri
249
Maiale
4,1 chilometri
862
Manzo
15,8 chilometri
3360
Quindi produrre 225 grammi di carne di manzo porta al rilascio di una quantità di gas serra quasi 13 volte maggiore di quella rilasciata a seguito della produzione di una eguale quantità di pollo e addirittura 57 volte maggiore se consideriamo le patate.
Per fare un altro esempio produrre i 41 chili di carne bovina consumata annualmente dallo statunitense medio genera la stessa quantità di CO2 di un’automobile che percorra 3000 chilometri.
Bibliografia
Di Daniele N., Noce A., Vidiri M., Moriconi E., Marrone G., Annicchiarico-Petruzzelli M., D’Urso G., Tesauro M., Rovella V., De Lorenzo A. Impact of Mediterranean diet on metabolic syndrome, cancer and longevity. Oncotarget 2017;8:8947-8979. doi:10.18632/oncotarget
Estruch R., Ros E., Salas-Salvadó J., et al. Primary prevention of cardiovascular disease with a Mediterranean Diet. N Engl J Med 2013;368:1279-1290. doi:10.1056/NEJMoa1200303
Friel S., Dangour A.D., Garnett T., Lock K., Chalabi Z., Roberts I., Butler A., Butler C.D., Waage J., McMichael A.J. and Haines A. Public health benefits of strategies to reduce greenhouse-gas emissions: food and agriculture. Lancet 2009;374:2016-2025. doi:10.1016/S0140-6736(09)61753-0
Giugliano D. and Esposito K. Mediterranean Diet and Cardiovascular Health. Annals NY Acad Sci 2005;1056(1):253-260. doi:10.1196/annals.1352.012
Giugliano D. and Esposito K. Mediterranean diet and metabolic diseases. Curr Opin Lipidol 2008;19:63-68. doi:10.1097/MOL.0b013e3282f2fa4d
Keys A. Mediterranean diet and public health: personal reflections. Am J Clin Nutr 1995;61:1321S-1323S doi:10.1093/ajcn/61.6.1321S
Keys A., Aravanis C., Blackburn H., Buzina R., Djordjevic B.S., Dontas A.S., Fidanza F., Karvonen M.J., Kimura N., Menotti A., Mohacek I., Nedeljkovic S., Puddu V., Punsar S., Taylor H.L., Van Buchem F.S.P. Seven Countries: A Multivariate Analysis of Death and Coronary Heart Disease. Harvard University Press, Cambridge, Harvard University Press, ISBN: 0-674-80237-3, 1980. 381 pp.
Martínez-González M.Á., de la Fuente-Arrillaga C., Nunez-Cordoba J.M., Basterra-Gortari F.J., Beunza J.J., Vazquez Z., Benito S., Tortosa A., Bes-Rastrollo M. Adherence to Mediterranean diet and risk of developing diabetes: prospective cohort study. BMJ 2008;336:1348-1351. doi:10.11bmj.39561.501007.BE
Martín-Peláez S., Fito M., Castaner O. Mediterranean diet effects on type 2 diabetes prevention, disease progression, and related mechanisms. A review. Nutrients 2020;12(8):2236. doi:10.3390/nu12082236
Mentella M.C., Scaldaferri F., Ricci C., Gasbarrini A., Miggiano G.A.D. Cancer and Mediterranean Diet: a review. Nutrients 2019; 11(9):2059. doi:10.3390/nu11092059
Nestle M. Mediterranean diets: historical and research overview. Am J Clin Nutr 1995;61:1313S-1320S doi:10.1093/ajcn/61.6.1313S
Samieri C., Okereke O.I., E. Devore E.E. and Grodstein F. Long-term adherence to the Mediterranean Diet is associated with overall cognitive status, but not cognitive decline, in women. J Nutr 2013;143:493-499. doi:10.3945/jn.112.169896
Sánchez-Villegas A., Delgado-Rodríguez M., Alonso A., Schlatter J., Lahortiga F., Serra Majem L., Martínez-González M.A. Association of the Mediterranean Dietary pattern with the incidence of depression: The Seguimiento Universidad de Navarra/University of Navarra Follow-up (SUN) Cohort. Arch Gen Psychiatry. 2009;66:1090-1098 doi:10.1001/archgenpsychiatry.2009.129
Scarmeas N., Luchsinger J.A., Mayeux R. and Stern Y. Mediterranean diet and Alzheimer disease mortality. Neurology 2007;69(11):1084-1093 doi:10.1212/01.wnl.0000277320.50685.7c
Schröder H. Protective mechanisms of the Mediterranean diet in obesity and type 2 diabetes. J Nutr Biochem 2007;18:149-160. doi:10.1016/j.jnutbio.2006.05.006
Sofi F., Cesari F., Abbate R., Gensini G.F. and Casini A. Adherence to Mediterranean diet and health status: meta-analysis. BMJ 2008;337:a1344. doi:10.1136/bmj.a1344
Subak S. Global environmental costs of beef production. Ecol. Econom. 1999;30:79-91. doi:10.1016/S0921-8009(98)00100-1
Trichopoulou A., Bamia C. and Trichopoulos D. Anatomy of health effects of Mediterranean diet: Greek EPIC prospective cohort study. BMJ 2009;338:b2337. doi:10.1136/bmj.b2337
Trichopoulou A., Costacou T., Bamia C., Trichopoulos D. Adherence to a Mediterranean Diet and Survival in a Greek Population. N Engl J Med 2003;348:2599-2608. doi:10.1056/NEJMoa025039
Il tratto gastrointestinale dell’uomo è una delle più feroci e competitive nicchie ecologiche presenti in natura. Vi si ritrovano virus, eucarioti, batteri, e una sola specie di Archeobatteri, Methanobrevibacter smithii.
I batteri variano in proporzione e quantità lungo tutto il tratto gastrointestinale. La presenza maggiore si ha nel colon, con oltre 400 specie diverse appartenenti a 9 fila o divisioni, ed è a questi che ci si riferirà parlando di microbiota intestinale, che a sua volta è parte del più ampio microbiota umano. Di seguito, l’elenco dei phyla suddetti e di alcuni tra i loro generi maggiormente rappresentati.
Actinobacteria, Gram-positivi; Bifidobacterium, Collinsella, Eggerthella e Propionibacterium.
Bacteroidetes, Gram-negativi; oltre 20 generi tra cui Bacteroides, Prevotella e Corynebacterium.
Cyanobacteria, Gram-negativi.
Firmicutes, Gram-positivi; almeno 250 generi tra cui Mycoplasma, Bacillus, Clostridium, Dorea, Faecalibacterium, Ruminococcus, Eubacterium, Staphylococcus, Streptococcus, Lactobacillus, Lactococcus,Enterococcus, Sporobacter e Roseburia.
Fusobacteria, Gram-negativi; Sneathia.
Lentisphaerae, Gram-negativi.
Proteobacteria, Gram-negativi; Escherichia, Klebsiella, Shigella, Salmonella, Citrobacter, Helicobacter e Serratia.
Spirochaeates, Gram-negativi.
Verrucomicrobia, Gram-negativi.
La presenza nel colon di un piccolo sottoinsieme del mondo batterico, 9 phyla sui 30 esistenti nel dominio Bacteria, è il risultato di una forte pressione selettiva che nel corso dell’evoluzione ha agito sia sui colonizzatori microbici, selezionando organismi adattati eccezionalmente bene a questo ambiente e in grado di dominare il processo di colonizzazione, che sulla nicchia intestinale. E tuttavia, ciascun individuo possiede nel proprio intestino una comunità batterica unica.
Nonostante la grande variabilità esistente sia riguardo ai taxa presenti che tra gli individui, è stato proposto, ma non da tutti accettato, che nella maggior parte dei soggetti adulti il microbiota intestinale, nella sua componente batterica, possa essere classificato in varianti o “enterotipi” sulla base del rapporto tra l’abbondanza di Bacteroides e Prevotella. Questo sembra indicare che esista un numero limitato di stati simbiotici ben bilanciati, che potrebbe rispondere in maniera differente a fattori quali la dieta, l’età, la genetica e l’assunzione di farmaci.
L’intestino degli adulti ospita un’ampia e varia comunità di virus a DNA e RNA, formata da circa 2000 genotipi differenti, dove però non se ne individua uno dominante, considerando che il virus più abbondante rappresenta solo circa il 6% dell’intera comunità, al contrario di ciò che accade nei neonati dove il genotipo più abbondante rappresenta oltre il 40% dell’intera comunità. La maggior parte dei virus a DNA sono batteriofagi o fagi, ossia virus che infettano i batteri, che sono anche l’entità biologica più abbondante presente sulla terra, con un una popolazione stimata di circa 1031 unità, mentre la maggior parte di quelli a RNA sono virus vegetali.
Influenze sulla composizione e sviluppo del microbiota intestinale
La comunità batterica intestinale è regolata da diversi fattori, molti dei quali sono di seguito elencati.
La dieta
La dieta dell’ospite sembra essere il fattore più importante, a partire dal primo alimento assunto, il latte materno.
Sebbene considerato sterile, il latte materno contiene un ricco microbiota formato da oltre 700 specie, dominato da stafilococchi, streptococchi, bifidobatteri e batteri lattici. Dunque, nei i bambini allattati al seno rappresenta una fonte importante per la colonizzazione dell’intestino, ed è stato suggerito che questa modalità di colonizzazione giochi un ruolo cruciale per la salute, in quanto, tra le altre funzioni potrebbe proteggere il neonato dalle infezioni e contribuire alla maturazione del sistema immunitario. Il latte materno influenza il microbiota intestinale anche indirettamente, grazie alla presenza di oligosaccaridi con attività prebiotica che stimolano la crescita di gruppi batterici specifici quali stafilococchi e bifidobatteri.
Anche uno studio che ha confrontato il microbiota intestinale di bambini europei e africani, rispettivamente da Firenze e un villaggio rurale del Burkina Faso, di età compresa tra 1 e 6 anni, ha messo in evidenza il ruolo decisivo della dieta rispetto ad altre variabili quali il clima, la geografia, l’igiene e i servizi sanitari; è stata inoltre osservata un’assenza di differenze significative nell’espressione di geni chiave nel regolare la funzione immune, che suggerisce quindi una similarità funzionale tra i due gruppi. Infatti i bambini di entrambe i gruppi, fintanto che sono allattati al seno, presentano un microbiota intestinale con caratteristiche molto simili, ricco in Actinobacteria, principalmente Bifidobacterium.
La successiva introduzione di una dieta solida differente nei due gruppi, di tipo Occidentale negli europei e dunque ricca in grassi e proteine animali, povera in proteine animali ma ricca in carboidrati complessi nei bambini africani, porta a una differenziazione del rapporto Firmicutes/Bacteroidetes nei due gruppi. Nei bambini europei erano più abbondanti i Gram-positivi, principalmente Firmicutes, rispetto ai Gram-negativi, mentre nei bambini africani prevalevano i Gram-negativi, principalmente Bacteroidetes, rispetto ai Gram-positivi.
E la dieta a lungo termine è associata in modo molto stretto alla ripartizione in enterotipi. E’ stato infatti osservato che:
una dieta ricca in grassi e proteine animali, dunque di tipo occidentale, porta a un microbiota intestinale dominato da taxa dell’enterotipo Bacteroides;
una dieta ricca in carboidrati, tipica delle società agricole, vede la prevalenza dell’enterotipo Prevotella.
Analoghi risultati sono emersi dallo studio sopracitato sui bambini. Negli europei, il microbiota intestinale era dominato da taxa tipici dell’enterotipo Bacteroides, mentre in quelli del Burkina Faso, dominano taxa dell’enterotipo Prevotella.
Con cambiamenti a breve termine della dieta, 10 giorni, quali il passaggio da una povera in grassi e ricca in fibre a una ricca in grassi e povera in fibre e viceversa, sono stati osservati cambiamenti nella composizione del microbioma, già dopo 24 ore, ma nessuno scambio stabile nella suddivisione in enterotipi. E questo rimarca come per un cambiamento dell’enterotipo del microbiota intestinale sia necessaria una dieta a lungo termine.
Modifiche a carico della dieta si traducono anche in cambiamenti a carico del viroma intestinale, che si sposta verso un nuovo stato, ossia si osservano alterazioni delle proporzioni delle popolazioni preesistenti, verso il quale convergono individui che seguano la stessa dieta.
Il pH, sali biliari ed enzimi digestivi
Lo stomaco, a causa del pH estremamente acido del suo contenuto, è un ambiente ostile per i batteri, che non sono presenti in numero elevato, circa 102-103 cellule batteriche/grammo di tessuto. Oltre a Helicobacter pylori, capace di causare gastriti e ulcere gastriche, sono presenti anche batteri del genere Lactobacillus.
Dal duodeno si osserva un incremento nel numero di unità, 104-105 cellule batteriche/grammo di tessuto; e quantità simili si ritrovano nel digiuno e nelle prime parti dell’ileo. Il numero contenuto di microorganismi presente nell’intestino tenue è dovuto all’ambiente inospitale conseguente al fatto che nel tratto discendente del duodeno è presente l’apertura dell’ampolla di Vater dalla quale viene riversata la bile e il succo pancreatico ossia sali biliari ed enzimi pancreatici, entrambe in grado di causare danni ai microrganismi presenti.
Nella porzione terminale dell’ileo, dove l’attività dei sali biliari e degli enzimi pancreatici è meno intensa, la conta batterica è di circa 107 cellule batteriche/grammo di tessuto, fino ad arrivare nel colon a valori pari a 1012-1014 cellule batteriche/grammo di tessuto, tanto che le feci sono costituite per il 40% da batteri.
La distribuzione di batteri lungo l’intestino è strategica. Nel duodeno e nel digiuno la quantità di nutrienti ancora disponibile è molto più alta rispetto a quella presente nell’ileo terminale, dove sono rimasti acqua, fibre, ed elettroliti. Dunque non è un problema trovare nell’ileo terminale, e ancor più nel colon, un numero elevato di batteri. Il problema sarebbe trovarli in numero eccessivo nel duodeno, digiuno e prima parte dell’ileo; ed esiste una condizione patologia, definita sindrome da sovracrescita batterica nel tenue o SIBO, acronimo dell’inglese small intestinal bacterial overgrowth, nella quale il numero di batteri nel tenue aumenta di circa 10-15 volte, il che li mette in condizione di poter competere con l’ospite per i nutrienti e dare origine a disturbi gastrointestinali quali ad esempio diarrea.
La posizione geografica e le conseguenti differenze riguardo lo stile di vita, di alimentazione, di religione ecc.
E’ stato ad esempio osservata una sorta di gradiente geografico nel microbiota dei neonati europei, con un numero più elevato di specie di Bifidobacterium e alcune di Clostridium nei bambini delle zone settentrionali, mentre in quelli delle zone meridionali è stata trovata una maggiore abbondanza di Bacteroides, Lactobacillus ed Eubacterium.
La modalità di nascita.
L’assetto genetico dell’ospite.
Lo stato di salute, anche della madre nel corso della gravidanza.
Ad esempio, in pazienti con malattia infiammatoria intestinale o IBD, acronimo dell’inglese inflammatory bowel disease, risulta depleta Faecalibacterium prausnitzii, una tra le specie produttrici di acido butirrico, un’importante fonte di energia per le cellule intestinali, e dotata anche di un’azione antiinfiammatoria in vitro e nei topi, mentre si osserva un aumento nel numero di Escherichia coli aderenti.
L’assunzione di antibiotici.
Le infezioni batteriche e i predatori.
Le batteriocine, ossia proteine dotate di attività antibatterica, e i batteriofagi.
Questi ultimi sono una forza importante nel controllo dell’abbondanza e composizione del microbiota intestinale. In particolare potrebbero avere un ruolo di primo piano nel corso della colonizzazione dell’intestino del neonato, infettando via via gli ospiti dominanti e creando così l’opportunità per un altro ceppo di divenire abbondante. Questo modello di dinamiche preda-predatore, definito “kill the winner”, suggerisce che le fioriture di determinate specie batteriche porterebbero a fioriture dei loro corrispondenti fagi, seguite da riduzioni dell’abbondanza di entrambe. Di conseguenza il genotipo fagico più abbondante non sarà lo stesso in momenti differenti. E sebbene alcune sequenze virali presenti nell’intestino del neonato siano stabili nel corso dei primi tre mesi di vita, sono state osservate drammatiche variazioni nella composizione complessiva della comunità virale fecale tra la prima e la seconda settimana. Infine, anche la comunità batterica nel corso di questo periodo è estremamente dinamica.
La competizione per lo spazio e i nutrienti.
Composizione nel corso della vita
Lo sviluppo dell’ecosistema microbico intestinale è un evento complesso e cruciale nella vita dell’uomo, altamente variabile da individuo a individuo, e influenzato dai fattori visti in precedenza.
Sviluppo e Modificazioni della Flora Batterica Intestinale
Nell’utero materno i bambini sono considerati sterili, e dunque soggetti al momento del parto alla colonizzazione da parte dei microbi, grazie anche al fatto di nascere con una tolleranza immunitaria “insegnata” dalla madre.
Tuttavia alcuni lavori stanno evidenziando la presenza di batteri nel tessuto placentare, nel sangue del cordone ombelicale, nelle membrane fetali e nel liquido amniotico di neonati sani senza indicazioni di infezioni o infiammazioni. E ad esempio, il meconio di neonati prematuri nati da madri sane contiene un microbiota specifico, con i Firmicutes come phylum principale e predominanza di stafilococchi, mentre nelle prime feci i più abbondanti sono i Proteobacteria, in particolare specie quali Escherichia coli, Klebsiella pneumoniae, Serratia marcescens, ma anche gli enterococchi.
Nota: il meconio è privo di particelle virali rilevabili.
Sembra che sia i batteri vaginali che quelli intestinali possano avere accesso al feto, anche se attraverso vie differenti: per via ascendente i primi, tramite le cellule dendritiche del sistema immunitario i secondi. Dunque potrebbe esistere anche un microbiota fetale.
Al momento del parto si verifica la colonizzazione ad opera di un piccolo inoculo di origine materna, formato in genere da aerobi obbligati e facoltativi (inizialmente l’intestino contiene ossigeno), poi sostituiti da anaerobi obbligati, i batteri tipici presenti nell’età adulta, cui i primi colonizzatori hanno preparato l’ambiente idoneo.
Inoltre è presente un basso numero di taxa differenti, con dominio relativo dei phyla Proteobacteria e Actinobacteria, che rimane tale anche durante il primo mese di vita, ma non nei successivi dove si verifica un grande aumento della variabilità, parallelo a quello di nuove varianti genetiche. Si ritiene che l’esposizione microbica iniziale sia importante nel definire le “traiettorie” che porteranno agli ecosistemi adulti. Infine, le comunità iniziali possono fungere da fonte diretta di batteri protettivi o patogeni molto presto nel corso della vita.
Nei bambini nati con parto naturale le più importanti fonti di inoculo sono il microbiota vaginale e fecale della madre. Infatti i neonati ospitano comunità microbiche dominate dai specie dei generi Lactobacillus, i più abbondanti nel microbiota vaginale e anche nell’intestino neonatale nei primi giorni, Bifidobacterium, Prevotella, o Sneathia. E sembra probabile che gli anaerobi, quali i membri dei phyla Bacteroidetes e Firmicutes, non potendo vivere all’esterno dei loro ospiti, facciano affidamento allo stretto contatto tra genitore e figli per la trasmissione. Infine la trasmissione degli anerobi stretti, data la presenza nell’intestino del neonato di ossigeno, potrebbe avvenire in seguito per mezzo di spore e non direttamente al momento del parto.
In caso di parto cesareo, i primi batteri incontrati sono quelli della pelle e dell’ambiente ospedaliero, e i neonati ospitano un microbiota dominato da specie appartenenti ai generi Corynebacterium, Staphylococcus e Propionibacterium, con una conta batterica intestinale più bassa e una minore diversità nelle prime settimane di vita rispetto ai bimbi nati con parto naturale.
Ulteriore prova che avvalora l’ipotesi di una trasmissione verticale è il fatto che esista una somiglianza tra il microbiota intestinale presente nel meconio, e campioni prelevati dai possibili siti di contaminazione. Queste “firme” materne non persistono indefinitamente, ma vengono sostituite da altre popolazioni microbiche entro il primo anno di vita.
Fonti successive di inoculo sono la bocca e la pelle dei parenti, oggetti, animali, ma sembra essere il latte materno ad avere il ruolo primario nel determinare la successione microbica a livello intestinale.
La variazione e la diversità presente invece tra i bambini rifletterebbe l’individualità di queste esposizioni accidentali.
Nota: il tipo di parto sembra influenzare anche le funzioni immunologiche nel corso del primo anno di vita, forse attraverso l’influenza esercitata sullo sviluppo del microbiota intestinale. I bambini nati con il cesareo avrebbero:
una più bassa conta di cellule batteriche nei campioni di feci a un mese dalla nascita, dovuta soprattutto alla più elevata presenza di Bifidobacterium nei bambini nati con parto naturale;
un più alto numero di cellule secretrici anticorpi, IgA, IgG e IgM, il che potrebbe riflettere un’eccessiva esposizione agli antigeni che riuscirebbero ad attraversare la più vulnerabile barriera intestinale.
Entro giorni dalla nascita si viene a creare una fiorente comunità la cui popolazione è molto più variabile in composizione e meno stabile nel tempo rispetto a quella dell’adulto, e che ben presto supererà in numero quella delle cellule del neonato stesso, evolvendo secondo un pattern temporale notevolmente variabile da individuo a individuo.
I virus, assenti alla nascita, dalla fine della prima settimana di vita raggiungono un numero di circa le 108 unità/grammo di peso fresco di feci, e quindi rappresentano una componente abbondante e dinamica del microbiota intestinale in via di sviluppo, ma con una diversità estremamente bassa, al pari di quella batterica. La comunità virale è dominata dai fagi, che probabilmente influenzano anche la diversità e abbondanza dei batteri concomitanti come visto in precedenza. La sorgente iniziale di virus non è nota; tra le possibili fonti ci sono ovviamente quelle materne e ambientali. D’altro canto i primi virus potrebbero essere il risultato dell’induzione dei profagi della “neonata” flora batterica, ipotesi avvalorata dall’osservazione che oltre il 25% delle sequenze fagiche sembrano essere molto simili a quelle dei fagi che infettano batterici come Lactobacillus, Lactococcus, Enterococcus, e Streptococcus, abbondanti nel latte materno.
Dal termine del primo mese di vita si ritiene che la fase iniziale di rapida acquisizione di microbi seguente alla nascita sia terminata.
Dal punto di vista tassonomico nei bambini di un mese i microrganismi più abbondanti sembrano essere quelli appartenenti ai generi Bacteroides ed Escherichia, mentre i Bifidobacterium compaiono e crescono fino a dominare, assieme a Ruminococcus, il tratto gastrointestinale del bambino allattato al seno tra il primo e l’undicesimo mese di vita.
I bifidobatteri, come Bifidobacterium longum subspecies infantis, sono:
strettamente correlati all’allattamento al seno;
tra i meglio caratterizzati fra i batteri commensali benefici;
probiotici, ossia microrganismi in grado di apportare effetti benefici sulla salute dell’ospite.
La loro abbondanza conferisce benefici anche attraverso un’esclusione competitiva con cui sono di ostacolo alla colonizzazione da parte di patogeni. E infatti Escherichia e Bacteroides possono divenire preponderanti se i Bifidobacterium non colonizzano in modo adeguato.
Al contrario, nei bambini allattati artificialmente batteri dei generi Escherichia, come E. coli, Clostridium, come C. difficile, Bacteroides, come B. fragilis e Lactobacillus sono presenti in modo significativamente maggiore rispetto a quanto osservato nei bambini allattati al seno.
Anche se prima dello svezzamento la dieta del neonato allattato al seno è piuttosto costante, il suo microbiota non lo è altrettanto. Si osservano infatti ampie fluttuazioni nell’abbondanza dei taxa batterici, con differenze tra soggetti che riguardano anche i pattern temporali di variazione. Le variazioni osservate potrebbero essere dovute a eventi casuali di colonizzazione, malattie, assunzione di antibiotici, cambiamenti nel comportamento dell’ospite o ad altri aspetti legati allo stile di vita, come anche a differenze nelle risposte immunitarie ai microbi colonizzatori. Ma come questi fattori contribuiscano a plasmare il microbiota intestinale del bambino non è ancora chiaro.
Sembra che anche il viroma si modifichi molto rapidamente dopo la nascita, in quanto la maggior parte delle sequenze virali presenti nella prima settimana di vita non si ritrova dopo la seconda, e il repertorio si espande rapidamente in diversità e numero nel corso dei primi tre mesi. E questo contrasta con la stabilità che si osserva nell’adulto dove il 95% delle sequenze si conserva nel tempo.
In condizioni normali, verso la fine del primo anno di vita, il neonato è stato esposto a una dieta complessa per un periodo significativo e dovrebbe aver sviluppato una comunità microbica con caratteristiche analoghe a quelle del microbiota intestinale adulto, quali:
una composizione più stabile, filogeneticamente più complessa, e progressivamente più simile tra soggetti diversi;
una preponderanze di Bacteroidetes e Firmicutes, una comune presenza di Verrucomicrobia e un’abbondanza molto bassa di Proteobacteria;
un aumento del carico batterico come dei livelli di acidi grassi a catena corta o SCFA, principalmente l’acido acetico, l’acido propionico e l’acido butirrico, nelle feci;
un aumento dei geni associati all’utilizzo dei carboidrati, alla biosintesi delle vitamine e alla degradazione degli xenobiotici.
Interessante notare che il significativo turn-over di taxa che si verifica dalla nascita al termine del primo anno è accompagnato da una notevole costanza nelle capacità funzionali complessive.
Verso la fine del primo anno di vita anche i colonizzatori virali sono oramai stati sostituiti da virus specifici del bambino.
Verso i due anni e mezzo di vita il microbiota intestinale raggiunge un suo equilibrio, somigliando in modo completo a quello dell’adulto.
La selezione dei batteri più altamente adattati è conseguenza di vari fattori.
Il passaggio a una dieta solida.
Una maggiore idoneità all’ambiente intestinale dei taxa che tipicamente dominano il microbiota colonico adulto rispetto agli opportunisti iniziali.
I cambiamenti progressivi nell’ambiente intestinale, dovuti ai cambiamenti legati allo sviluppo della mucosa stessa.
Gli effetti del microbiota medesimo.
Quindi i primi 2-3 anni di vita sono il periodo più critico in cui intervenire al fine di ottenere il miglior microbiota possibile così da ottimizzare la crescita e lo sviluppo del bambino.
Tutto questo, partendo da un inizio caotico, porta alla costituzione dell’ecosistema intestinale tipico del giovane adulto, che è relativamente stabile nel tempo, comprese le componenti virale, eucariotica e gli Archeobatteri, sino alla vecchiaia, dominato almeno nella popolazione occidentale da specie dei phyla Firmicutes, che rappresentano circa il 60% della comunità batterica intestinale, Bacteroidetes e Actinobacteria, principalmente con il genere Bifidobacterium, ognuno circa il 10% della comunità batterica, seguiti da Proteobacteria e Verrucomicrobia. I generi Bacteroides, Clostridium, Faecalibacterium, Ruminococcus ed Eubacterium, costituiscono, assieme a Methanobrevibacter smithii, la grande maggioranza della comunità batterica intestinale dell’adulto.
Risultati contrastanti con i dati esposti stanno emergendo dall’analisi di popolazioni ad esempio di villaggi rurali dell’Africa, come visto sopra.
E il microbiota intestinale è sufficientemente simile tra gli individui da permettere l’individuazione di un nucleo microbiomico condiviso.
La stabilità e resilienza è però soggetta a numerose variabili tra cui, come detto, la dieta sembra essere una delle più importanti, variabili che quindi dovranno essere mantenute costanti, o nel caso della malattie evitate, ad esempio attraverso le vaccinazioni, al fine di mantenere la stabilità del microbiota. Va comunque sottolineato che stabilità e resilienza potrebbero essere dannose nel caso in cui la comunità intestinale dominante sia patogena.
Negli anziani, come già visto nei neonati, il microbiota intestinale va incontro a sostanziali cambiamenti. In uno studio condotto in Irlanda su 161 persone sane di 65 o più anni, nella maggior parte dei soggetti il microbiota intestinale è distinto da quello degli adulti più giovani, con una composizione che sembra essere dominato dai phyla Bacteroidetes, i principali, seguiti dai Firmicutes, con percentuali quasi inverse rispetto a quelle trovate negli adulti più giovani, anche se sono state osservate notevoli variazioni tra i soggetti. E tra i generi più abbondanti si trovano Faecalibacterium, che rappresenta circa il 6% dei 15 generi principali, seguiti dalle specie dei generi Ruminococcus, Roseburia e Bifidobacterium, quest’ultimo intorno allo 0,4%.
Anche la variabilità nella composizione delle comunità è più grande rispetto ai giovani adulti, il che potrebbe essere dovuto alla maggior morbilità associata con l’età, e quindi il successivo utilizzo di medicinali, nonché a variazioni della dieta.
Bibliografia
Breitbart M., Haynes M., Kelley S., Angly F., Edwards R.A., Felts B., Mahaffy J.M., Mueller J., Nulton J., Rayhawk S., Rodriguez-Brito B., Salamon P., Rohwer F. Viral diversity and dynamics in an infant gut. Res Microbiol 2008;159:367-373. doi:10.1016/j.resmic.2008.04.006
Claesson M.J., Cusack S., O’Sullivan O., Greene-Diniz R., de Weerd H., Flannery E., Marchesi J.R., Falush D., Dinan T., Fitzgerald G., et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc Natl Acad Sci USA 2011;108(Suppl 1);4586-4591. doi:10.1073/pnas.1000097107
Clemente J.C., Ursell L.K., Wegener Parfrey L., and Knight R. The impact of the gut microbiota on human health: an integrative view. Cell 2012;148:1258-1270. doi:10.1016/j.cell.2012.01.035
De Filippo c., Cavalieri D., Di Paola M., Ramazzotti M., Poullet J.B., Massart S., Collini S., Pieraccini G., and Lionetti P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci 2010;107(33):14691-14696. doi:10.1073/pnas.1005963107
Dominguez-Bello M.G., Costello E.K., Contreras M., Magris M., Hidalgo G., Fierer N., and Knight R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci 2010;107:11971-11975. doi:10.1073/pnas.1002601107
Fernández L., Langa S., Martín V., Maldonado A., Jiménez E., Martín R., Rodríguez J.M. The human milk microbiota: origin and potential roles in health and disease. Pharmacol Res 2013;69(1):1-10. doi:10.1073/pnas.1002601107
Huurre A., Kalliomäki M., Rautava S., Rinne M., Salminen S., and Isolauri E. Mode of delivery-effects on gut microbiota and humoral immunity. Neonatology 2008;93:236-240. doi:10.1159/000111102
Koenig J.E., Spor A., Scalfone N., Fricker A.D., Stombaugh J., Knight R., Angenent L.T., and Ley R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci 2011;108(1):4578-4585. doi:10.1073/pnas.1000081107
Ley R.E., Peterson D.A., and Gordon J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006;124(4):837-848. doi:10.1016/j.cell.2006.02.017
Minot S., Sinha R., Chen J., Li H., Keilbaugh S.A., Wu G.D., Lewis J.D., and Bushman F.D. The human gut virome: inter-individual variation and dynamic response to diet. Genome Res 2011;21:1616-1625. doi:10.1101/gr.122705.111
Moreno-Indias I.M., Cardona F., Tinahones F.J. and Queipo-Ortuño M.I. Impact of the gut microbiota on the development of obesity and type 2 diabetes mellitus. Front Microbiol 2014;5(190):1-10. doi:10.3389/fmicb.2014.00190
Newburg D.S. & Morelli L. Human milk and infant intestinal mucosal glycans guide succession of the neonatal intestinal microbiota. Pediatr Res 2015;77:115-120. doi:10.1038/pr.2014.178
Palmer C., Bik E.M., DiGiulio D.B., Relman D.A., and Brown P.O. Development of the human infant intestinal microbiota. PLoS Biol 2007;5(7):e177. doi:10.1371/journal.pbio.0050177
Rodrıguez J.M., Murphy K., Stanton C., Ross R.P., I. Kober O.I., Juge N., Avershina E., Rudi K., Narbad A., Jenmalm M.C., Marchesi J.R. and Collado M.C. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb Ecol Health Dis 2015;26:26050. doi:10.3402/mehd.v26.26050
Wu G.D., Chen J., Hoffmann C., Bittinger K., Chen Y.Y., Keilbaugh S.A., Bewtra M., Knights D., Walters W.A., Knight R., et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011;334:105-108. doi:10.1126/science.1208344
E’ noto da quasi un secolo che gli esseri umani ospitano un ecosistema microbico, definito microbiota umano, straordinariamente denso e diversificato, formato da un numero di virus e cellule molto superiore a quello che compone il corpo umano, e che rappresenta dall’uno al tre per cento del peso corporeo.
I geni che i microrganismi componenti il microbiota umano codificano, che sono circa 1000 volte più numerosi rispetto a quelli del nostro genoma, formano il microbioma umano.
I microrganismi colonizzano tutte le superfici del corpo esposte all’ambiente. Distinte comunità microbiche sono infatti presenti sulla pelle, nella vagina, nelle vie aeree, e lungo tutto il tubo digerente, a partire dalla bocca, passando attraverso lo stomaco sino a raggiungere le parti terminali dell’intestino.
II microbiota umano è costituito da organismi provenienti da tutti i taxa. Vi si ritrovano infatti batteri, virus, archeobatteri ed eucarioti.
Batteri
I batteri sono presenti con almeno cento trilioni (1014) di cellule, un numero dieci volte maggiore rispetto a quelle che compongono il corpo umano.
Si ritrovano per la maggior parte a livello del tratto intestinale, dove, con concentrazioni sino a 1012-1014/grammo di tessuto, formano uno degli habitat più densamente popolati esistenti sulla terra. In questa sede sono particolarmente abbondanti membri dei phyla Firmicutes, ma anche di Bacteroidetes e Actinobacteria.
Nota: le comunità microbiche di una data sede si “somigliano” tra di loro molto di più di quanto le stesse non somiglino a quelle presenti in altri siti dello stesso soggetto; ad esempio, le comunità delle vie aeree superiori sono molto più simili tra individui differenti che non a quelle della pelle o dell’intestino di uno stesso soggetto.
Virus
I virus sono in assoluto i componenti più numerosi essendo presenti con quadrilioni di unità. I genomi di tutti i virus ospitati costituiscono il viroma umano.
In passato i virus e gli eucarioti del microbiota umano sono stati studiati focalizzandoci sui microrganismi patogeni, ma negli ultimi anni l’attenzione si è spostata anche sui numerosissimi membri non patogeni di questi gruppi. E riguardo ai virus, molte delle sequenze geniche trovate sono nuove, il che suggerisce che ci sia ancora molto da conoscere sul viroma umano.
Infine, al pari dei batteri, anche per i virus esiste una notevole variabilità interpersonale.
Archeobatteri
Gli Archeobatteri o archei, nome scientifico Archaea, sono rappresentati in particolare dai microrganismi appartenenti all’ordine dei Methanobacteriales. Tra questi Methanobrevibacter smithii predomina nell’intestino umano, rappresentando sino al 10% di tutti gli anaerobi.
Eucarioti
Anche gli eucarioti sono presenti, e tra i primi ad essere identificati probabilmente ci sono i parassiti del genere Giardia e Entamoeba. Ma vi è anche una grande abbondanza e diversità di funghi, appartenenti ad esempio a generi quali Candida, Penicillium, Aspergillus, Hemispora, Fusarium, Geotrichum, Cryptococcus, Hormodendrum, Saccharomyces e Blastocystis.
Candida albicans
Funzione del microbiota umano
Talvolta definito “l’organo dimenticato”, il microbiota umano, e in particolare la sua componente batterica intestinale, svolge numerose ed importanti funzioni che possono portare a benefici nutrizionali, immunologici, e di sviluppo, ma può anche essere causa di malattie per l’ospite.
Di seguito alcuni esempi.
E’ coinvolto nello sviluppo del sistema gastrointestinale, come dimostrato da esperimenti condotti su animali germ-free nei quali, ad esempio, lo spessore della mucosa intestinale è più sottile rispetto a quella di animali colonizzati, dunque più facilmente soggetto ad insulti che ne causino la rottura.
Concorre all’estrazione dell’energia dai nutrienti, grazie alla sua capacità di fermentare carboidrati per noi indigeribili; inoltre promuove l’assorbimento dei monosaccaridi ed il deposito dell’energia ricavata. Tutto ciò con molta probabilità ha rappresentato una forza evolutiva molto forte che ha giocato a favore del fatto che questi batteri siano diventati nostri simbionti.
Concorre al mantenimento del pH acido della pelle e del contenuto del colon.
E’ coinvolto nel metabolismo degli xenobiotici e di molti polifenoli.
Migliora l’assorbimento di acqua e sali minerali (ferro, calcio e magnesio) nel colon.
Aumenta la velocità di transito intestinale, più lenta negli animali germ-free.
Ha un ruolo importante nella resistenza alla colonizzazione da parte di microrganismi patogeni, in particolare nella vagina e nell’intestino.
E’ coinvolto nella biosintesi di isoprenoidi e vitamine attraverso la via del metileritritolo fosfato.
Stimola l’angiogenesi.
Interagisce con il sistema immunitario, fornendo segnali per promuovere la maturazione delle cellule immunitarie ed il normale sviluppo delle funzioni immuni. E questo è forse l’effetto più importante derivante dalla simbiosi tra uomo e microrganismi. Esperimenti condotti su animali germ-free hanno infatti evidenziato quanto segue.
I macrofagi, le cellule che hanno il compito di fagocitare i patogeni e poi presentarne gli antigeni al sistema immunitario, sono presenti in scarsissima quantità rispetto all’intestino colonizzato, e se messi in presenza di batteri non riescono a trovarli e quindi a fagocitarli, a differenza dei macrofagi estratti da un intestino colonizzato.
Manca la flogosi cronica aspecifica, una condizione normale del nostro intestino dovuta alla presenza di un tessuto immunitario molto sviluppato e allenato, proprio grazie alla presenza dei batteri nel lume intestinale (e anche di ciò che mangiamo).
Cambiamenti nella sua composizione possono contribuire allo sviluppo di obesità e sindrome metabolica.
Protegge dallo sviluppo del diabete di tipo I.
Molte malattie, sia del bambino che dell’adulto, tra cui il tumore dello stomaco e del colon, i linfomi del tessuto linfoide associato alla mucosa, l’enterocolite necrotizzante, che è un’importante causa di morbilità e mortalità nei prematuri, o le malattie croniche intestinali, sono e altre appaiono essere collegate al microbiota intestinale.
Sembra quindi molto probabile che l’organismo umano rappresenti un superorganismo frutto di tanti anni di evoluzione, composto oltre che dalle proprie cellule e capacità metaboliche e fisiologiche che ne derivano, anche da un altro organo aggiunto, il microbiota.
Commensali e patogeni
Sulla base dei rapporti che stabiliscono con l’ospite, i differenti microrganismi componenti il microbiota umano possono essere quindi suddivisi in due categorie: commensali e patogeni.
I commensali non causano danno all’ospite, con cui anzi instaurano una simbiosi di tipo mutualistico che in genere porta benefici ad entrambe.
I patogeni sono al contrario in grado di causare malattie, ma fortunatamente rappresentano una minima percentuale della nostra flora microbica. Si tratta di microrganismi che stabiliscono una simbiosi con l’ospite traendone beneficio a suo svantaggio.
In genere causano malattia se si verifica:
un “cambio di sede”, ossia se si spostano dalla loro normale nicchia, ad esempio l’intestino, ad un’altra impropria, quale la vagina o la vescica, come nel caso del fungo Candida albicans presente normalmente, ma in piccolissima quantità, nell’intestino;
un indebolimento delle difese immunitarie dell’ospite, come dopo un trapianto o comunque una terapia immunosoppressiva.
Human Microbiome Project
La componente batterica del microbiota umano è l’oggetto di vari studi tra i quali un progetto molto ampio partito nel 2008 chiamato “Human Microbiome Project”, che ne analizza il microbioma associato a vari habitat del corpo, quali pelle, bocca, naso, vagina ed intestino, in una popolazione sana di 242 adulti.
Questi studi hanno evidenziato l’esistenza di una grande variabilità nella composizione del microbiota umano, con i gemelli che condividono meno del 50% dei loro taxa batterici a livello di specie, ed una percentuale anche minore riguardo i virus.
I fattori che modellano la composizione delle comunità microbiche iniziano ad essere compresi. Ad esempio le caratteristiche genetiche dell’ospite hanno un ruolo importante nel creare e plasmare le comunità batteriche presenti, anche se questo non è vero per quelle virali. E studi di metagenomica hanno evidenziato che, nonostante la grande variabilità interpersonale nella composizione delle comunità microbiche, esiste un ampio nucleo condiviso di geni codificante vie del segnale e metaboliche. Sembra cioè che l’assemblaggio e la struttura delle comunità microbiche non avvenga in base alle specie quanto al gruppo più funzionale di geni.
E da questo ne deriverebbe che stati di malattia di queste comunità possano essere meglio identificati da distribuzione atipiche di classi di geni funzionali.
Effetti degli antibiotici
l microbiota di un individuo adulto sano è generalmente stabile nel tempo. Tuttavia la sua composizione può essere alterata ad opera di fattori esterni come l’urbanizzazione, i viaggi , le modifiche della dieta, ma soprattutto l’uso di antibiotici a largo spettro.
Gli antibiotici hanno un profondo effetto.
Vi è una riduzione a lungo termine nella diversità batterica.
I taxa colpiti variano da individuo ad individuo, e posso essere interessati fino ad un terzo di quelli presenti.
Alcuni taxa non recuperano anche dopo 6 mesi dal trattamento.
Una volta che la comunità batterica si è rimodellata dopo il trattamento con il farmaco, si osserva una ridotta resistenza alla colonizzazione. Ciò permette a microbi estranei e/o patogeni in grado di crescere più dei commensali di causare cambiamenti permanenti nella struttura del microbiota umano, come anche malattie sia acute, ad esempio la pericolosa colite pseudomembranosa, che croniche, come si sospetta per l’asma a seguito dell’uso ed abuso di antibiotici nell’infanzia.
Inoltre il loro ripetuto uso sembra aumentare la riserva di geni per la resistenza agli antibiotici nel nostro microbioma. A supporto di questa ipotesi in alcune nazioni europee è stata osservata una riduzione nel numero dei patogeni antibiotico-resistenti a seguito di un calo nel numero degli antibiotici prescritti.
Infine non va sottovalutato il fatto che la microflora batterica intestinale è implicata in molte trasformazioni chimiche, per cui una sua alterazione potrebbe avere implicazioni nello sviluppo del cancro come dell’obesità.
Tuttavia, riguardo all’uso degli antibiotici va sottolineato che se abbiamo un’aspettativa di vita molto superiore al passato è anche perché non moriamo più di malattie infettive!
Bibliografia
Burke C., Steinberg P., Rusch D., Kjelleberg S., and Thomas T. Bacterial community assembly based on functional genes rather than species. Proc Natl Acad Sci USA 2011;108:14288-14293. doi:10.1073/pnas.1101591108
Clemente J.C., Ursell L.K., Wegener Parfrey L., and Knight R. The impact of the gut microbiota on human health: an integrative view. Cell 2012;148:1258-1270. doi:10.1016/j.cell.2012.01.035
Gill S.R., Pop M., Deboy R.T., Eckburg P.B., Turnbaugh P.J., Samuel B.S., Gordon J.I., Relman D.A., Fraser-Liggett C.M., and Nelson K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006;312:1355-1359. doi:10.1126/science.1124234
Palmer C., Bik E.M., DiGiulio D.B., Relman D.A., and Brown P.O. Development of the human infant intestinal microbiota. PLoS Biol 2007;5(7):e177. doi:10.1371/journal.pbio.0050177
Turnbaugh P.J., Gordon J.I. The core gut microbiome, energy balance and obesity. J Physiol 2009;587:4153-4158. doi:10.1113/jphysiol.2009.174136
Zhang, T., Breitbart, M., Lee, W., Run, J.-Q., Wei, C., Soh, S., Hibberd, M., Liu, E., Rohwer, F., Ruan, Y. Prevalence of plant viruses in the RNA viral community of human feces. PLoS Biol 2006;4(1):e3. doi:10.1371/journal.pbio.0040003
L’olio di oliva è un grasso alimentare ottenuto dalla spremitura delle olive, i frutti dell’olivo (Olea europaea).
È un elemento fondamentale della dieta mediterranea e un prodotto tipico dell’area mediterranea, con una storia millenaria e un grande valore economico.
Dal punto di vista chimico nell’olio di oliva è possibile individuare, sulla base del comportamento in presenza di soluzioni fortemente alcaline e calore, due frazioni:
la frazione saponificabile, che rappresenta il 98-99% del peso totale, ed è formata da lipidi che, nelle condizioni suddette, formano saponi;
la frazione insaponificabile, che costituisce il restante 1-2% del peso, e che nelle condizioni suddette non forma saponi.
E’ composta da acidi grassi saturi e acidi grassi insaturi, quasi per intero legati al glicerolo a formare trigliceridi o triacilgliceroli. In misura molto minore si ritrovano anche digliceridi o diacilgliceroli, monogliceridi o monoacilgliceroli, e acidi grassi liberi.
L’acido oleico è l’acido grasso più abbondante dell’olio di oliva, con una concentrazione che, in base alle norme stabilite dall’International Olive Oil Council (IOOC) deve essere compresa tra il 55% e 83% del totale degli acidi grassi.
L’acido linoleico è l’acido grasso polinsaturo più abbondante, con una concentrazione che deve essere comprese tra il 2,5% e il 21%. A causa del suo elevato grado di insaturazione è soggetto facilmente a ossidazione; questo significa che un olio che ne contenga quantità elevate irrancidisce con più facilità, e dunque può essere conservato per un tempo minore rispetto a un olio che ne contenga meno.
Nella cucina mediterranea l’olio di oliva è la principale fonte di grassi: dunque l’acido oleico, tra gli acidi grassi monoinsaturi, e il linoleico, tra i polinsaturi, sono i principali acidi grassi presenti.
L’acido alfa-linolenico deve essere presente in quantità molto bassa, secondo gli standard IOOC ≤1%. Si tratta di un acido grasso polinsaturo della famiglia degli acidi grassi polinsaturi omega-3, che può apportare benefici dal punto di vista nutrizionale, ma, a causa della forte insaturazione, al pari dell’acido linoleico, è molto suscettibile a ossidazione, e quindi favorisce l’irrancidimento dell’olio che lo contiene.
La presenza di acidi grassi che non dovrebbero essere presenti, o che dovrebbero essere presenti in percentuali diverse rispetto a quelle osservate, sono indici di adulterazione dell’olio, che è stato “tagliato” con altri oli di semi. A questo riguardo particolare attenzione viene rivolta agli acidi miristico, arachidico, beenico, lignocerico, gadoleico e alfa-linolenico, le cui percentuali ammissibili sono specificate dal regolamento IOOC.
La composizione in acidi grassi è influenzata da diversi fattori.
Il clima.
La latitudine.
La zona di produzione.
Gli oli prodotti in Italia, Spagna e Grecia sono ricchi in acido oleico e poveri in acido linoleico e palmitico, mentre l’olio tunisino è ricco in acido linoleico e palmitico e più povero in acido oleico. Dunque, gli oli d’oliva possono essere suddivisi in due gruppi:
uno con elevato contenuto di acido oleico e basso di acido linoleico e palmitico;
l’altro con un elevato contenuto in acido linoleico e palmitico e basso di acido oleico.
Il tipo di cultivar.
Il grado di maturazione delle olive al momento dell’estrazione dell’olio.
Va notato che l’acido oleico è formato per primo nel frutto, e i dati sembrano indicare una forte rapporto antagonistico tra l’acido oleico e gli acidi palmitico, palmitoleico e linoleico.
Trigliceridi
Come detto, gli acidi grassi dell’olio di oliva sono quasi per intero legati a formare molecole di trigliceridi.
In piccola percentuale si ritrovano anche legati in molecole di digliceridi, monogliceridi, oppure in forma libera.
Numerazione Stereospecifica o sn
Durante la biosintesi dei trigliceridi, grazie alla presenza di specifici enzimi, solo circa il 2% delle molecole del glicerolo legherà in posizione sn-2 una molecola di acido palmitico; anche la percentuale di acido stearico in posizione 2 è molto bassa. Per la maggior parte la posizione sn-2 sarà invece occupata da acido oleico.
Al contrario, se si considerano oli che abbiano subito processi di esterificazione artificiali, viene a mancare la specificità dell’azione enzimatica, per cui la percentuale di acido palmitico in posizione sn-2 aumenta in modo significativo.
Nota: sn- è l’acronimo dell’inglese sterospecific numbering, che significa numerazione sterospecifica.
Tra i trigliceridi presenti in proporzioni più significative si ritrovano:
OOO (40-59%);
POO (12-20%);
OOL (12,5-20%);
POL (5,5-7%);
SOO (3-7%).
Si ritrovano anche quantità minori di POP, POS, OLnL, OLnO, PLL, PLnO.
La trilinoleina (LLL) è un trigliceride contenente tre molecole di acido linoleico. Un suo basso livello è indice di buona qualità dell’olio.
Non sono stati osservati trigliceridi con soli acidi grassi saturi e neppure contenenti solo residui di acido alfa-linolenico.
Digliceridi e monogliceridi
La loro presenza è dovuta sia ad una incompleta sintesi dei trigliceridi che a parziali reazioni di idrolisi dei trigliceridi stessi.
La concentrazione dei digliceridi nell’olio di oliva vergine varia tra 1% e il 2,8%. Nell’olio fresco prevalgono gli 1,2-digliceridi, rappresentando oltre l’80% dei digliceridi. Nel corso della conservazione dell’olio si verifica una isomerizzazione con un progressivo incremento della forma 1,3, più stabile, che dopo circa 10 mesi diviene la forma principale. Dunque il rapporto tra 1,2- e 1,3-digliceridi può essere utilizzato come indicatore del grado di invecchiamento dell’olio.
I monogliceridi sono presenti in quantità inferiori rispetto ai digliceridi, meno dello 0,25%, con gli 1-monogliceridi ben più abbondanti dei 2-monogliceridi.
Frazioni insaponificabile
E’ composta da un elevato numero di molecole differenti molto importanti dal punto di vista nutrizionale, in quanto contribuiscono in modo considerevole a quelli che sono gli effetti salutari dell’olio di oliva. Inoltre sono responsabili della stabilità e del sapore dell’olio di oliva, e sono utilizzati anche per determinarne una eventuale adulterazione con altri oli.
Vi si ritrovano tocoferoli, steroli, polifenoli, pigmenti, idrocarburi, alcol aromatici e alifatici, acidi triterpenici, cere, e composti minori.
Il loro contenuto è influenzato da fattori in parte analoghi a quelli visti per la composizione in acidi grassi, quali:
il tipo di cultivar;
il grado di maturazione dell’oliva;
la zona di produzione;
l’anno di raccolta e le modalità di raccolta delle olive;
il tempo di stoccaggio delle olive;
il tipo di lavorazione per l’estrazione dell’olio;
le condizioni di conservazione dell’olio.
Da sottolineare che negli oli di oliva raffinati molti di questi composti non sono presenti in quanto rimossi durante la lavorazione.
Polifenoli
I polifenoli rappresentano il 18-37% della frazione insaponificabile.
Sono un gruppo molto eterogeneo di molecole con proprietà sia nutrizionali che organolettiche, come l’oleuropeina e l’idrossitirosolo che danno all’olio un gusto amaro e pungente.
Per una loro più ampia trattazione si rimanda all’articolo: Polifenoli dell’olio di oliva.
Idrocarburi
Rappresentano il 30-50% della frazione insaponificabile.
I principali sono lo squalene e il beta-carotene.
Lo squalene, isolato per la prima volta dal fegato di squalo, è il principale componente della frazione insaponificabile, e rappresenta oltre il 90% della frazione idrocarburica; la sua concentrazione oscilla tra i 200 e i 7500 mg/kg di olio.
E’ un intermedio nella via di sintesi del nucleo a 4 anelli degli steroidi. Sembra essere responsabile, almeno in parte, degli effetti benefici dell’olio di oliva.
Nella frazione idrocarburica dell’olio vergine di oliva, oltre allo squalene, si ritrovano anche idrocarburi diterpenici e triterpenici, poliolefine isoprenoidi e n-paraffine.
Il beta-carotene agisce sia come antiossidante, proteggendo l’olio durante la conservazione, che come colorante.
Steroli
Sono un’importante gruppo di lipidi dell’olio di oliva che hanno:
diversi effetti salutari;
si correlano con la qualità dell’olio;
sono ampiamente utilizzati per testarne la genuinità.
A questo riguardo c’è da sottolineare il fatto che sono molecole specie specifiche, per cui trovare ad esempio nell’olio di oliva elevate concentrazioni di brassicasterolo, uno sterolo tipico delle Brassicaceae, come la colza, indica una sua adulterazione con olio di canola.
Si individuano 4 classi di steroli: steroli comuni, 4-metilsteroli, alcol triterpenici, e dialcol triterpenici. Il loro livello nell’olio di oliva oscilla tra 1000 mg/kg, il valore minimo richiesto dagli standard IOOC, e 2000 mg/kg. I valori più bassi si osservano negli oli raffinati, dove il processo di raffinazione può causare perdite anche del 25%.
Steroli comuni o 4-alfa-desmetilsteroli
Gli steroli comuni sono presenti per la maggior parte in forma libera ed esterificata, sebbene siano stati rinvenuti anche come lipoproteine e steril glucosidi.
I principali sono il beta-sitosterolo, che rappresenta dal 75 al 90% del totale degli steroli, il Δ5-avenasterolo, tra il 5 e il 20%, e il campesterolo, 4%.Altre molecole presenti in quantità inferiori o tracce sono ad esempio il stigmasterolo, 2%, colesterolo, brassicasterolo, ergosterolo.
4-Metilsteroli
Sono intermedi nella biosintesi degli steroli, e si ritrovano sia in forma libera che esterificata.
Sono presenti in piccole quantità, molto minori rispetto a quelle degli steroli comuni e degli alcol triterpenici, oscillando tra 50 e 360 mg/kg. Le molecole principali sono l’obtusifoliolo, il gramisterolo, cicloeucalenolo, e il citrostadienolo.
Alcol triterpenici o 4,4-dimetilsteroli
Sono una classe molto complessa di steroli, e si ritrovano sia in forma libera che esterificata.
Sono presenti in quantità variabili tra 350 e 1500 mg/kg.
I principali sono β-amirina, butirrospermolo, 24 metilen cicloartanolo, e cicloartenolo; altre molecole presenti in quantità inferiori sono ad esempio il ciclobranolo, il ciclosadolo, dammaradienolo, e il germanicolo.
Dialcol triterpenici
I principali dialcol triterpenici dell’olio di oliva sono l’eritrodiolo e l’uvaolo.
L’eritrodiolo è presente sia in forma libera che esterificata, in quantità totale, nell’olio vergine di oliva, oscillante tra i 19 e i 69 mg/kg, mentre la forma libera è in genere inferiore a 50 mg/kg.
Tocoferoli
I tocoferoli rappresentano il 2-3% della frazione insaponificabile, e includono la vitamina E.
Degli otto vitameri della vitamina E, nell’olio di oliva vergine l’alfa-tocoferolo costituisce circa il 90% dei tocoferoli. E’ presente in forma libera e in quantità molto variabile, ma comunque in media superiore ai 100 mg/kg d’olio. Grazie alle sue proprietà antiossidanti in vivo, la sua abbondante presenza è un fattore molto positivo per la salute. La concentrazione dell’alfa-tocoferolo sembra essere correlata agli alti livelli di clorofille e alla concomitante richiesta di disattivazione dell’ossigeno singoletto.
Beta-tocoferolo, delta-tocoferolo e gamma-tocoferolo sono in genere presenti in basse quantità.
Pigmenti
In questo gruppo si ritrova le clorofille e i carotenoidi.
Le clorofille sono presenti come feofitine, in particolare nella forma alfa, ossia una clorofilla cui è stato rimosso il magnesio, e conferiscono all’olio di oliva il caratteristico colore verde. Sono molecole fotosensibilizzanti per cui contribuiscono alla foto-ossidazione dell’olio stesso.
I principali carotenoidi sono il beta-carotene e la luteina; sono presenti anche xantofille, quali anteraxantina, beta-criptoxantina, luteoxantina, mutatoxantina, neoxantina, violaxantina.
Il colore dell’olio di oliva è la risultante della presenza di clorofille e carotenoidi e delle loro tonalità verde e giallo. La loro presenza è strettamente correlata.
Acidi triterpenici
Gli acidi triterpenici sono componenti importanti dell’oliva dove sono presenti in tracce.
I principali acidi triterpenici presenti nell’olio vergine di oliva sono l’acido oleanolico e l’acido maslinico, ritrovandosi nella sansa da cui possono essere estratti in piccola quantità durante la lavorazione.
Alcol alifatici e aromatici
I più importanti sono gli alcol grassi e gli alcol diterpenici.
Gli alcol alifatici hanno un numero di atomi di carbonio pari, compreso tra 20 e 30, e per la maggior parte sono localizzati all’interno del nocciolo, da dove sono parzialmente estratti attraverso processi meccanici.
Alcol grassi
Sono alcol saturi lineari con più di 16 atomi di carbonio.
Sono presenti nell’olio vergine di oliva in quantità di solito non superiori a 250 mg/kg. Possono essere presenti sia in forma libera che esterificata.
I principali sono il docosanolo, tetracosanolo, esacosanolo, e l’octacosanolo, mentre le molecole con un numero dispari di atomi di carbonio sono presenti in tracce. Tetracosanolo ed esacosanolo sono quelli presenti in quantità maggiore.
Le cere, componenti minori dell’olio di oliva, sono esteri degli alcol grassi con acidi grassi. Le principali cere presenti sono esteri dell’acido palmitico e oleico. Possono essere utilizzate come criterio per differenziare i vari tipi di olio di oliva; ad esempio nell’olio vergine ed extravergine di oliva devono essere presenti in quantità inferiori a 150 mg/kg, in accordo con gli standard stabiliti dalla IOOC.
Alcol diterpenici
Fitolo e geranilgeraniolo sono due alcol diterpenici aciclici, presenti sia in forma libera che esterificata. Tra gli esteri nella frazione cerosa dell’olio extravergine di oliva si ritrovano quelli con l’oleato, eicosanoato, eicosenoato, docosanoato e tetracosanoato, principalmente in forma di fitil derivati.
Composti volatili
Nell’olio di oliva sono stati identificati oltre 280 composti volatili, tra cui idrocarburi, i più abbondanti, alcol, aldeidi, chetoni, esteri, acidi, eteri e altri. Solo circa 70 di questi composti sono presenti a livelli superiori a quella che è la soglia di percezione oltre la quale le molecole contribuiscono all’aroma dell’olio vergine di oliva.
Componenti minori
Tra i componenti minori si ritrovano fosfolipidi, di cui i principali sono la fosfatidilcolina, fosfatidiletanolamina, fosfatidilinositolo e fosfatidilserina.
Possono essere presenti, negli oli non filtrati, quantità minime di proteine.
Bibliografia
Gunstone F.D. Vegetable oils in food technology: composition, properties and uses. 2th Edition. Wiley J. & Sons, Inc., Publication, 2011
Caponio F., Bilancia M.T., Pasqualone A., Sikorska E., Gomes T. Influence of the exposure to light on extra virgin olive oil quality during storage. Eur Food Res Technol 2005;221:92-98. doi:10.1007/s00217-004-1126-8
Servili M., Sordini B., Esposto S., Urbani S., Veneziani G., Di Maio I., Selvaggini R. and Taticchi A. Biological activities of phenolic compounds of extra virgin olive oil. Antioxidants 2014;3:1-23. doi:10.3390/antiox3010001
Il glutine non è una semplice proteina, ma è una miscela composta da proteine dei cereali, per circa l’80% del suo peso secco, ad esempio gliadine e glutenine per il frumento, lipidi, 5-7%, amido, 5-10%, acqua, 5-8%, e sostanze minerali, <2%.
Si forma quando componenti naturalmente presenti nel chicco del cereale, la cariosside, e nella farina derivata, si uniscono tra di loro, in ambiente acquoso e sotto l’azione di sollecitazioni meccaniche, ossia durante la formazione dell’impasto.
Il termine è associato anche alla famiglia di proteine che causa problemi ai soggetti affetti da celiachia.
Isolato per la prima volta dal chimico italiano Jacopo Bartolomeo Beccari nel 1745 dalla farina di frumento, può essere estratto dall’impasto lavando lo stesso in modo delicato sotto acqua corrente: una volta allontanato l’amido, le albumine e le globuline, tutti solubili in acqua, rimane una massa appiccicosa ed elastica, appunto il glutine, termine che deriva dal latino gluten, che significa colla.
grano duro (Triticum durum), da cui si ottengono semole e semolati per pasta alimentare secca;
grano tenero (Triticum aestivum), da cui si ottengono farine per pane, paste fresche e prodotti da forno;
segale (Secale cereale);
orzo (Hordeum vulgare);
farro, nelle tre specie:
piccolo o monococco (Triticun monococcum);
medio (Triticum dicoccum Schrank);
rande o granfarro o spelta (Triticum spelta)
grano khorasan, di cui il Kamut® ne è una varietà;
triticale, che è un ibrido tra la segale e il grano tenero (× Triticosecale Wittmack);
bulgur o grano spezzato, che è grano duro integrale germogliato e successivamente lavorato;
seitan, che non è un cereale ma un derivato del frumento, da alcuni definito anche “bistecca di glutine”.
Dato che la maggior parte del glutine assunto con l’alimentazione proviene dalle farine di frumento, di cui se ne raccoglie circa 700 milioni di tonnellate annue, che rappresentano circa il 30% della produzione mondiale dei cereali, la discussione seguente sarà incentrata sul glutine di frumento, e in particolare sulle sue proteine.
Nota: il termine glutine viene anche utilizzato per indicare il residuo proteico che rimane dopo aver allontanato l’amido e le proteine solubili dall’impasto ottenuto con farina di mais o granturco: questo “glutine di mais” è tuttavia “funzionalmente” differente rispetto a quello ottenuto dal frumento.
Le proteine dei cereali
Lo studio delle proteine dei grani, come visto, ebbe inizio con il lavoro di Beccari.
In seguito, nel 1924, quindi ben 150 anni dopo, l’inglese Osborne T.B., che a ragione può essere considerato il padre della chimica delle proteine vegetali, ne sviluppò una classificazione sulla base della loro solubilità in vari solventi.
La classificazione, ancora in uso, suddivide le proteine vegetali in 4 famiglie.
Albumine, solubili in acqua.
Globuline, solubili in soluzioni saline, come l’avenalina dell’avena.
Prolamine, solubili in soluzione alcolica al 70%, ma non in acqua o alcol assoluto.
Comprendono:
gliadine del frumento;
zeina del mais;
avenina dell’avena;
ordeina dell’orzo;
secalina della segale.
Sono le responsabili dell’effetto tossico del glutine per il celiaco.
Gluteline, insolubili in acqua e soluzioni saline neutre, ma solubili in soluzioni acide e basiche.
Comprendono le glutenine del frumento.
Cereale
Albumine
Globuline
Prolamine*
Gluteline**
Frumento
9
5
40
46
Mais
4
2
55
39
Orzo
13
12
52
23
Avena
11
56
9
23
Riso
5
10
5
80
* Nel frumento gliadine
** Nel frumento glutenine
Albumine e globuline sono proteine citoplasmatiche, spesso di natura enzimatica, ricche di aminoacidi essenziali, quali lisina, triptofano e metionina. Si ritrovano nell’aleurone e nell’embrione della cariosside.
Prolamine e gluteline sono le proteine di riserva dei cereali. Sono ricche in asparagina, glutammina, arginina e prolina, ma molto povere in lisina, triptofano e metionina. Si ritrovano nell’endosperma, e rappresentano la grande maggioranza delle proteine presenti nel frumento, mais, orzo, avena e segale.
Sebbene la classificazione di Osborne sia ancora ampiamente utilizzata, sarebbe più corretto suddividere le proteine dei grani in tre gruppi: di riserva, strutturali e metaboliche, e con funzioni difensive.
Le proteine del glutine di frumento
Nel frumento le proteine rappresentano il 10-14% del peso della cariosside, mentre circa l’80% del peso è costituito da carboidrati.
Seguendo la classificazione di Osborne, il 15-20% delle proteine sono rappresentate dalla albumine e globuline, mentre il restante 80-85%, è costituito da prolamine e gluteline, rispettivamente gliadine, 30-40%, e glutenine, 40-50%. Quindi, e a differenza delle prolamine e gluteline degli altri cereali, gliadine e gluteine sono presenti in quantità simili, circa il 40%.
Proteine del Grano
Gliadine e glutenine hanno una notevole importanza dal punto di vista tecnologico. Perché?
Le proteine appartenenti alle due classi sono insolubili in acqua, e nell’impasto, dunque in un ambiente ricco d’acqua, si legano tra loro attraverso legami quali:
legami covalenti, ossia ponti disolfuro;
legami non covalenti, quali interazioni idrofobiche, forze di van der Waals, legami idrogeno e legami ionici.
Grazie alla formazione di questi legami intermolecolari, si crea un reticolo tridimensionale, che intrappola i granuli di amido e le bolle di anidride carbonica che si formano durante la lievitazione, e conferisce resistenza ed elasticità all’impasto di farina e acqua, due proprietà del glutine ampiamente sfruttate industrialmente.
Nella dieta abituale della popolazione europea adulta, e in particolare di quella italiana che è molto ricca di derivati del frumento, gliadine e glutenine sono le proteine maggiormente rappresentate, circa 15 g al giorno. Che significa? Che la dieta gluten-free è una dieta che impegna sia sotto l’aspetto psicologico che sociale la persona affetta da celiachia.
Nota: i lipidi componenti il glutine sono strettamente legati alle zone idrofobiche di gliadine e glutenine e, rispetto a quanto è possibile fare con la farina originale, sono rimossi con maggiore difficoltà (il contenuto in lipidi del glutine dipende dal contenuto in lipidi della farina da cui è stato ottenuto).
Gliadine: estensibilità e viscosità
Le gliadine sono prolamine idrofobiche monomeriche, cioè formate da una sola subunità, di natura globulare e con basso peso molecolare. Sulla base della mobilità elettroforetica in condizioni di basso pH, sono state suddivise nei seguenti gruppi:
alfa/beta, e gamma, ricche di zolfo, contenendo residui di cisteina, coinvolti nella formazione di ponti disolfuro intramolecolari, e di metionina;
omega, povere di zolfo, data l’assenza o quasi di cisteina e metionina.
Hanno uno scarso valore nutrizionale e un’altissima tossicità per il celiaco per la presenza di particolari sequenza aminoacidiche nella struttura primaria, in particolare prolina-serina-glutammina-glutammina e glutammina-glutammina-glutammina-prolina.
Le gliadine si associano tra di loro e con le glutenine attraverso legami non covalenti; grazie a ciò, nella formazione dell’impasto agiscono come “plasticizzanti”. Infatti, a loro si deve la viscosità e l’estensibilità proprie del glutine, il cui reticolo tridimensionale proteico si può deformare permettendo l’aumento di volume della massa a seguito della produzione di gas con la lievitazione. Questa proprietà è importante nella panificazione.
Un loro eccesso comporta la formazione di un impasto assai estensibile.
Glutenine: elasticità e tenacità
Le glutenine sono proteine polimeriche, ossia formate da più subunità, di natura filamentosa legate insieme da ponti disolfuro intermolecolari. Dopo riduzione dei suddetti legami, tramite SDS-PAGE possono essere suddivise in due gruppi.
Glutenine a elevato peso molecolare o HMW, acronimo dell’inglese high molecolar weight.
Povere di zolfo, rappresentano circa il 12% del totale delle proteine del glutine. I legami non covalenti che si stabiliscono tra le subunità di questo gruppo sono responsabili dell’elasticità e tenacità delle network di proteine del glutine, ossia delle proprietà viscoelastiche del glutine stesso e quindi anche dell’impasto che lo contiene.
Glutenine a basso peso molecolare o LMW, acronimo dell’inglese low molecolar weight.
Ricche di zolfo (cisteina), formano ponti disolfuro tra di loro e con le subunità HMW, formando così un macropolimero di glutenina.
Le glutenine fanno si che l’impasto mantenga la sua forma durante gli stress meccanici, come l’impastamento, e non meccanici, come l’aumento di volume dovuto alla lievitazione e all’aumento di volume dei gas che intrappola in conseguenza del riscaldamento dovuto alla cottura, cui è sottoposto. Questa proprietà è importante nella pastificazione.
Se in eccesso, le glutenine portano alla formazione di un impasto forte e rigido.
Proprietà del glutine di frumento
Dal punto di vista nutrizionale le proteine che compongono il glutine non hanno un elevato valore biologico, essendo povere di lisina, un aminoacido essenziale. Dunque una dieta senza glutine non comporta alcuna carenza significativa di nutrienti importanti.
Di contro, il glutine ha un grande valore per l’industria alimentare: la matrice proteica tridimensionale derivante dalla combinazione in soluzione acquosa di gliadine e glutenine, conferisce proprietà viscoelastiche, ossia di estensibilità-viscosità ed elasticità-tenacità, all’impasto di cui fa parte, e quindi una struttura ben definita al pane, alla pasta, e in generale a tutti gli alimenti che si fanno con la farina di frumento.
Ha un alto grado di palatabilità.
Ha un elevato potere fermentante a livello dell’intestino tenue.
E’ un’esorfina: alcuni peptidi derivati dalla digestione delle proteine del glutine possono andare ad agire a livello del sistema nervoso centrale.
Cereali senza glutine
Di seguito una lista di cereali, cereali minori, e pseudocereali gluten-free utilizzati a fini alimentari.
Cereali
Mais o granturco (Zea mais)
Riso (Oryza sativa)
Cereali minori
Definiti “minori” non perché di scarsa importanza nutrizionale, quanto perché coltivati in piccole aree e in quantità inferiori rispetto a frumento, riso e mais.
Fonio (Digitaria exilis)
Miglio (Panicum miliaceum)
Panico (Panicum italicum)
Sorgo (Sorghum vulgare)
Teff (Eragrostis tef)
Teosinte; gruppo composto da 4 specie appartenenti al genere Zea. Sono piante che crescono in Messico (Sierra Madre), Guatemala e Venezuela.
Pseudocereali
Così definiti perché associano nella loro botanica e nei loro aspetti nutrizionali caratteristiche peculiari dei cereali e dei legumi, quindi di un’altra famiglia di piante.
Grano saraceno (Fagopyrum esculentum)
Quinoa (Chenopodium quinoa), uno pseudocereale con ottime proprietà nutritive, contenendo fibre, ferro, zinco e magnesio, che fa parte della famiglia delle Chenopodiaceee, come le barbabietole.
Manioca, anche nota come tapioca, yuca e cassava (Manihot utilissima). Coltivata principalmente nel sud del Sahara e nell’America del Sud, è una radice tubero commestibile da cui si origina la fecola di tapioca.
Nota: non sempre i prodotti naturalmente privi di glutine al momento della commercializzazione sono effettivamente gluten-free. Infatti per gli alimenti preparati a livello industriale ci può essere o l’utilizzo di derivati contenenti gliadina o una contaminazione nella filiera produttiva, e questo è ovviamente importante in quanto anche tracce di glutine nella dieta possono causare problemi al celiaco.
Avena e glutine
Discorso a parte merita l’avena (Avena sativa), che è tra i cereali concessi ai celiaci. Studi condotti negli ultimi anni hanno evidenziato che è tollerata dal celiaco, adulto e bambino, anche nel soggetto con dermatite erpetiforme. Ovviamente, deve essere certificata per l’assenza di glutine da contaminazione.
Bibliografia
Beccari J.B. De Frumento. De bononiensi scientiarum et artium instituto atque Academia Commentarii, II. 1745:Part I.,122-127
Bender D.A. “Benders’ dictionary of nutrition and food technology”. 8th Edition. Woodhead Publishing. Oxford, 2006
Berdanier C.D., Dwyer J., Feldman E.B. Handbook of nutrition and food. 2th Edition. CRC Press. Taylor & Francis Group, 2007
Phillips G.O., Williams P.A. Handbook of food proteins. 1th Edition. Woodhead Publishing, 2011
Shewry P.R. and Halford N.G. Cereal seed storage proteins: structures, properties and role in grain utilization. J Exp Bot 2002:53(370);947-958. doi:10.1093/jexbot/53.370.947
Yildiz F. Advances in food biochemistry. CRC Press, 2009
Nella corsa il runner va incontro a un consumo calorico e a una perdita di acqua e sali minerali che dipendono da diversi fattori quali la tecnica, il grado di allenamento, le condizioni ambientali e le caratteristiche fisiologiche proprie di ciascun atleta. Ne consegue che la loro conoscenza permette di impostare un’alimentazione adeguata sia nella fase di allenamento che in quella di recupero tra un allenamento e il successivo, con l’obbiettivo di ottimizzare la performance.
Di seguito verrà analizzato il consumo calorico del runner impegnato in allenamenti su diverse distanze, e, nel dettaglio la quantità di carboidrati, lipidi, e proteine ossidati per ricavare l’energia necessaria a sostenere il lavoro muscolare, e quali sono i sali minerali maggiormente persi con la sudorazione.
Nella corsa il consumo calorico è pari a 0,85-1,05 kcal per kg di peso corporeo al chilometro.
Il range è conseguenza del fatto che gli atleti dotati di un’ottima tecnica consumano meno energia rispetto a quelli con una tecnica meno raffinata.
Un runner di 70 kg avrà un consumo calorico al chilometro compreso tra:
70 x 0,85 x 1 = 59,5 kcal
e
70 x 1,05 x 1 = 73,5 kcal
Nella tabella sono mostrati i calcoli per derivare la spesa calorica sostenuta dall’atleta per correre 10, 20, 30, e 40 km.
Distanza
Consumo calorico
10 km
0,85 x 70 x 10 = 595 kcal
1,05 x 70 x 10 = 735 kcal
20 km
0,85 x 70 x 20 = 1190 kcal
1,05 x 70 x 20 = 1470 kcal
30 km
0,85 x 70 x 30 = 1785 kcal
1,05 x 70 x 30 = 2205 kcal
40 km
0.85 x 70 x 40 = 2380 kcal
1.05 x 70 x 40 = 2940 kcal
Nota: chi ha cominciato a correre da poco avrà spese maggiori di 1,05 Kcal per kg di peso corporeo al chilometro.
L’energia richiesta per sostenere il lavoro muscolare nella corsa deriva dall’ossidazione di carboidrati, lipidi, e proteine. Carboidrati e lipidi rappresentano la principale fonte di energia, e il loro utilizzo è influenzato dall’intensità dell’esercizio: al suo aumentare si riduce la percentuale dei lipidi ossidati mentre aumenta quella dei carboidrati, come riassunto di seguito.
Intensità
Carburante
<30% VO2max
Principalmente grassi
40-60% VO2max
Grassi e carboidrati egualmente
75% VO2max
Principalmente carboidrati
>80% VO2max
Circa 100% carboidrati
Nota: L’impossibilità di utilizzare il carburante metabolico adeguato può promuovere la fatica e portare al sovrallenamento.
Quindi per andature maggiori della soglia anerobica, l’ossidazione dei carboidrati può arrivare a soddisfare l’intera richiesta energetica. Nell’andatura tipica della maratona, i carboidrati forniscono il 60-70% dell’energia necessaria, mentre per andature inferiori la percentuale si riduce a valori inferiori al 50%.
Di seguito verrà analizzato il consumo di carboidrati, lipidi e proteine nell’allenamento, nel corso del quale il dispendio energetico è sostenuto per circa il 60% dai carboidrati, per circa il 40% dai lipidi, mentre la percentuale residua, compresa tra il 3% e il 5%, dalle proteine
Ossidazione dei carboidrati nell’allenamento
Per un runner di 70 kg, il consumo di carboidrati al chilometro sarà compreso tra:
(0,6 x 59,5) / 4 = 8,9 g/km
e
(0,6 x 73,5) / 4 = 11 g/km
Nota: un grammo di carboidrati apporta circa 4 kcal.
Nella tabella sono mostrati i calcoli per derivare il consumo di carboidrati per correre 10, 20, 30, e 40 km.
Distanza
Consumo di carboidrati
10 km
[(0,85 x 70 x 10) x 0,6] / 4 = 89 g
[(1,05 x 70 x 10) x 0,6] / 4 = 110 g
20 km
[(0,85 x 70 x 20) x 0,6] / 4 = 179 g
[(1,05 x 70 x 20) x 0,6] / 4 = 221 g
30 km
[(0,85 x 70 x 30) x 0,6] / 4 = 268 g
[(1,05 x 70 x 30) x 0,6] / 4 = 331 g
40 km
[(0,85 x 70 x 40) x 0,6] / 4 = 357 g
[(1,05 x 70 x 40) x 0,6] / 4 = 441 g
Ossidazione dei lipidi nell’allenamento
Con calcoli simili a quelli fatti per i carboidrati è possibile derivare il consumo di lipidi al chilometro, che sarà compreso tra:
(0,4 x 59,5)/9 = 2,6 g/km
e
(0,4 x 73,5)/9 = 3,3 g/km
Nota: un grammo di lipidi apporta circa 9 kcal.
Nella tabella sono mostrati i calcoli per derivare il consumo di lipidi per correre 10, 20, 30, e 40 km.
Distanza
Consumo di lipidi
10 km
[(0,85 x 70 x 10) x 0,4] / 9 = 26 g
[(1,05 x 70 x 10) x 0,4] / 9 = 33 g
20 km
[(0,85 x 70 x 20) x 0,4] / 9 = 53 g
[(1,05 x 70 x 20) x 0,4] / 9 = 65 g
30 km
[(0,85 x 70 x 30) x 0,4] / 9 = 79 g
[(1,05 x 70 x 30) x 0,4] / 9 = 98 g
40 km
[(0,85 x 70 x 40) x 0,4] / 9 = 106 g
[(1,05 x 70 x 40) x 0,4] / 9 = 131 g
Ossidazione delle proteine nell’allenamento
Il fabbisogno proteico giornaliero di un soggetto adulto è pari a 0,9 grammi per kg di peso corporeo, e, per un atleta di 70 kg corrisponde a:
70 x 0,9 = 63 g
Durante la corsa circa il 3-5% dell’energia consumata per sostenere il lavoro muscolare deriva dall’ossidazione delle proteine.
Nella tabella sono mostrati i calcoli per derivare la quantità di proteine ossidate quando l’atleta corre per 10, 20, 30 e 40 km, considerando che le proteine forniscano il 3% dell’energia necessaria..
Distanza
Consumo di proteine al 3%
10 km
[(0,85 x 70 x 10) x 0,03)] / 4 = 4,5 g
[(1,05 x 70 x 10) x 0,03)] / 4 = 5,5 g
20 km
[(0,85 x 70 x 20) x 0,03)] / 4 = 8,9 g
[(1,05 x 70 x 20) x 0,03)] / 4 = 11 g
30 km
[(0,85 x 70 x 30) x 0,03)] / 4 = 13,4 g
[(1,05 x 70 x 30) x 0,03)] / 4 = 16,5 g
40 km
[(0,85 x 70 x 40) x 0,03)] / 4 = 17,9 g
[(1,05 x 70 x 40) x 0,03)] / 4 = 22,1 g
Nota: un grammo di proteine apporta circa 4 kcal.
Considerando il dispendio energetico di 0,85 e 1,05 kcal per kg di peso corporeo al chilometro, il fabbisogno proteico medio aggiuntivo per chilo di peso corporeo per correre 10, 20, 30, e 40 chilometri, approssimato alla seconda cifra decimale, è pari a:
10 km: [(4,5 + 5,5) / 2] / 70 = 0,07 g
20 km: [(4,5 + 5,5) / 2] / 70 = 0,14 g
30 km: [(4,5 + 5,5) / 2] / 70 = 0,21 g
40 km: [(4,5 + 5,5) / 2] / 70 = 0,29 g
Infine, considerando anche il fabbisogno proteico giornaliero del soggetto adulto si ottiene il fabbisogno proteico complessivo di un atleta di 70 kg impegnato nelle quattro distanze:
10 km: 0,07 + 0,9 = 0,97 g
20 km: 0,14 + 0,9 = 1,04 g
30 km: 0,21 + 0,9 = 1,11 g
40 km: 0,29 + 0,9 = 1,19 g
Con calcoli analoghi ai precedenti si ottiene il fabbisogno proteico complessivo nel caso di un’ossidazione delle proteine pari al 5%.
10 km: 0,12 + 0,9 = 1,02 g
20 km: 0,24 + 0,9 = 1,14 g
30 km: 0,36 + 0,9 = 1,26 g
40 km: 0,48 + 0,9 = 1,38 g
Se si escludono gli atleti che si allenano tutti i giorni per 30 km o più, i valori ottenuti sono di poco superiori a 0,9 grammi per kg di peso corporeo.
In realtà il fabbisogno proteico giornaliero è leggermente superiore a quello calcolato in quanto una certa quantità di azoto, dunque di proteine si perde, oltre che con le urine, anche attraverso la sudorazione.
Perdita di acqua e sali minerali nella corsa
Le perdite di acqua dipendono dalla quantità di sudore che l’atleta produce, che a sua volta dipendente da:
temperatura e umidità dell’aria;
irraggiamento solare.
La perdita sarà tanto maggiore quanto più alti sono questi valori.
Va comunque sottolineato che il sudore è prodotto in quantità diverse da soggetto a soggetto.
Con la sudorazione i principali sali minerali persi sono:
il sodio e il cloro, circa 1 grammo per litro di sudore nell’atleta abituato ad allenarsi in condizioni ambientali che provocano una intensa sudorazione;
il potassio, in quantità corrispondente a circa il 15% del sodio perso;
il magnesio in quantità corrispondente a circa l’1% del sodio perso.
La quantità di sali persi è funzione del volume di sudore prodotto e aumenta negli atleti non acclimatati al caldo.
Nella tabella sono mostrati i valori, espressi in grammi/litro, dei minerali presenti nel sudore di atleti non acclimatati e acclimatati.
Atleti non acclimatati
Atleti acclimatati
Sodio
1,38
0,92
Cloro
1,5
1,00
Potassio
0,20
0,15
Magnesio
0,01
<0,01
Totale
3,09
2,08
Da quanto detto deriva che nel corso dell’attività fisica il minerale più utile da assumere è il sodio.
Dopo l’attività fisica il corridore, o chi suda molto, tende a mangiare più salato. Il fenomeno, scoperto nel corso di studi condotti su operai di fonderie, è conosciuto come fame selettiva. Per il potassio e il magnesio probabilmente non esiste la fame selettiva.
Bibliografia
Sawka M.N., Burke L.M., Eichner E.R., Maughan, R.J., Montain S.J., Stachenfeld N.S. American College of Sports Medicine position stand: exercise and fluid replacement. Med Sci Sport Exercise 2007;39(2):377-390. doi:10.1249/mss.0b013e31802ca597
Shirreffs S., Sawka M.N. Fluid and electrolyte needs for training, competition and recovery. J Sport Sci 2011;29:sup1, S39-S46. doi:10.1080/02640414.2011.614269
L’acido gamma-linolenico o GLA, un acido grasso essenziale appartenente al gruppo degli acidi grassi polinsaturi omega-6, come il suo precursore acido linoleico (il più abbondante acido grasso polinsaturo nell’epidermide, dove è coinvolto nel mantenimento della barriera epidermica all’acqua) svolge un ruolo importante nella fisiologia e fisiopatologia della pelle.
Studi condotti sugli esseri umani hanno rivelato che l’acido gamma-linolenico:
migliora l’umidità della pelle;
migliora la fermezza e la rugosità della pelle;
diminuisce la perdita d’acqua transepidermica (una delle anomalie della pelle negli animali sottoposti a carenza di acidi grassi essenziali).
PGE1
Utilizzando epidermide di cavia come un modello di epidermide umana (sono funzionalmente simili) è stato dimostrato che la supplementazione di animali con cibi ricchi di acido gamma-linolenico si traduce in una grande produzione nella pelle di PGE1 e 15-HETrE (come già dimostrato in a esperimenti in vitro).
Poiché queste molecole hanno entrambe proprietà anti-inflammatorie/anti-proliferative, la supplementazione con alimenti ricchi di acido gamma-linolenico può essere un’aggiunta alla terapia standard per i disturbi infiammatori/proliferativi della pelle.
Fonti principali di acido gamma-linolenico
Le principali fonti di acido gamma-linolenico sono gli oli di semi di:
ribes nero (da 15% al 19% degli acidi grassi totali);
enotera (dal 7% al 14% degli acidi grassi totali);
Pressione arteriosa e acido gamma-linolenico
La relazione tra assunzione di acidi grassi con la dieta e la pressione arteriosa provengono principalmente da studi condotti su ratti geneticamente modificati che spontaneamente sviluppano ipertensione (un modello animale comunemente usato per l’ipertensione umana.
In questi studi sono state osservate molte anomalie della membrana. Pertanto l’ipertensione nel “modello ratto” potrebbe essere correlata ai cambiamenti nel metabolismo degli acidi grassi polinsaturi a livello della membrana cellulare.
Riguardo agli acidi grassi polinsaturi, vari gruppi di ricerca hanno riportato che l’acido gamma-linolenico riduce la pressione arteriosa in ratti normali e geneticamente modificati (effetto maggiore) ed è stato ipotizzato che si verifichi attraverso l’interferenza con il sistema renina-angiotensina (che promuove la resistenza vascolare e la ritenzione renale), alterando le proprietà della membrana cellulare delle cellule muscolari lisce vascolari e quindi interferendo con l’azione dell’angiotensina II.
Un altro possibile meccanismo d’azione dell’acido gamma-linolenico per ridurre la pressione arteriosa potrebbe essere attraverso il suo metabolita acido diomo-gamma-linolenico, il quale può essere incorporato nei fosfolipidi della membrana delle cellule muscolari lisce vascolari, poi rilasciato dall’azione della fosfolipasi A2, e trasformato dalla COX-1 in PGE1 che induce il rilassamento della muscolatura liscia vascolare.
Artrite reumatoide e acido gamma-linolenico
In uno studio condotto da Leventhal e colleghi nel 1993 è stato dimostrato che l’assunzione di una elevata concentrazione di olio di borragine (circa 1400 mg/die di acido gamma-linolenico) per 24 settimane, ha portato a una riduzione clinicamente significativa dei segni e sintomi di attività dell’artrite reumatoide.
In uno studio successivo condotta da Zurier e colleghi nel 1996, l’assunzione di una dose più alta (circa 2,8 g/die di acido gamma-linolenico) per 6 mesi ha ridotto, in modo clinicamente rilevante, segni e sintomi dell’attività della malattia. I pazienti che sono rimasti per un anno sulla dose di 2,8 g/die nella dieta hanno mostrato un miglioramento continuato dei sintomi (l’uso di acido gamma-linolenico anche alla dose più alta, è ben tollerato, con minimi effetti collaterali).
Questi dati sottolineano che la quantità giornaliera e la durata dell’assunzione di acido gamma-linolenico con la dieta si correlano con l’efficacia clinica.
Bibliografia
Akoh C.C. and Min D.B. “Food lipids: chemistry, nutrition, and biotechnology” 3th ed. 2008
Chow Ching K. “Fatty acids in foods and their health implication” 3th ed. 2008
Fan Y.Y. and Chapkin R.S. Importance of dietary γ-linolenic acid in human health and nutrition. J Nutr 1998;128:1411-1414. doi:10.1093/jn/128.9.1411
Leventhal L.J., Boyce E.G. and Zurier R.B. Treatment of rheumatoid arthritis with gammalinolenic acid. Ann Intern Med 1993;119:867-873. doi:10.7326/0003-4819-119-9-199311010-00001
Miller C.C. and Ziboh V.A. Gammalinolenic acid-enriched diet alters cutaneous eicosanoids. Biochem Biophys Res Commun 1988;154:967-974. doi:10.1016/0006-291X(88)90234-3
Zurier R.B., Rossetti R.G., Jacobson E.W., DeMarco D.M., Liu N.Y., Temming J.E., White B.M. and Laposata M. Gamma-linolenic acid treatment of rheumatoid arthritis. A randomized, placebocontrolled trial. Arthritis Rheum 1996;39:1808-1817. doi:10.1002/art.1780391106
Molti studi hanno dimostrato una relazione diretta, dose-dipendente tra l’assunzione di alcool e la pressione sanguigna, in particolare per assunzioni superiori a due drink/die.
Tale relazione è indipendente da:
età;
apporto di sale;
obesità;
infine, persiste a prescindere dal tipo di bevanda.
Inoltre il forte consumo di bevande alcoliche per lunghi periodi di tempo è uno dei fattori predisponenti all’ipertensione; dal 5 al 7% dei casi della malattia sono dovuti a un eccessivo consumo di alcool.
Una meta-analisi di 15 studi clinici controllati randomizzati ha mostrato che la riduzione dell’assunzione di bevande alcoliche porta benefici terapeutici sia per gli ipertesi che i normotesi con riduzioni simili della pressione sistolica e diastolica (negli ipertesi la riduzione si verifica in poche settimane).
Assunzione di alcool e prevenzione dell’ipertensione
Le linee guida sulla prevenzione primaria dell’ipertensione raccomandano che il consumo di alcool (etanolo) nella maggior parte degli uomini, in assenza di controindicazioni, dovrebbe essere inferiore ai 28 g/die, limite entro il quale può ridurre il rischio di malattia cardiaca coronarica. Il consumo limitato a questi quantitativi deve essere ottenuto con l’assunzione di bevande a basso contenuto alcolico, preferibilmente ai pasti (bere quantità anche piccole o moderate al di fuori dei pasti aumenta la probabilità di sviluppare ipertensione): ciò significa non più di 680 ml di una birra normale o 280 ml di vino (12% vol), soprattutto in caso di ipertensione; per le donne e i soggetti esili il consumo dovrebbe essere dimezzato1.
Da evitare il consumo di bevande ad alta gradazione alcolica anche se il contenuto totale di etanolo non supera i 28 g/die.
Effetti dell’etanolo sulla pressione arteriosa
In ogni caso resta incertezza riguardo i benefici o i rischi attribuibili a un consumo di etanolo leggero/moderato sul rischio di ipertensione.
In uno studio pubblicato nell’aprile 2008 gli autori hanno esaminato l’associazione tra l’assunzione di etanolo e il rischio di sviluppare ipertensione in 28848 donne arruolate nel “Women’s Health Study” e 13455 uomini dal “Physicians’ Health Study”.
Lo studio conferma che un forte consumo di bevande alcoliche, superiore a 2 drink/die, aumenta il rischio di ipertensione sia negli uomini che nelle donne ma, sorprendentemente, ha rilevato che l’associazione tra un consumo leggero/moderato, fino a due drink/die, e il rischio di sviluppare l’ipertensione è diverso nei due sessi. Le donne hanno un rischio potenziale ridotto di ipertensione da un consumo di alcool leggero/moderato con un’associazione a forma di J2; gli uomini non hanno benefici da tale consumo ma anzi un aumento del rischio.
Tuttavia, le linee guida per la prevenzione primaria dell’ipertensione limitano il consumo di bevande alcoliche a meno di due drink/die negli uomini e meno di uno nei soggetti più esili e nelle donne.
1. Un drink standard contiene circa 14 g di etanolo, vale a dire una birra normale da 340 ml, 140 ml di vino (12% vol), o 42 ml di bevande alcoliche distillate, che sono comunque sconsigliate.
2. Molti studi hanno mostrato una relazione a forma di J tra l’assunzione di etanolo e la pressione sanguigna. I bevitori leggeri (non più di 28 g di etanolo/die) hanno una pressione più bassa rispetto agli astemi; invece chi consuma più di 28 g di etanolo/die ha una pressione più alta dei non bevitori. Quindi l’alcool potrebbe essere un vasodilatatore a basse dosi e un vasocostrittore a dosi maggiori.
Sesso H.D., Cook N.R., Buring J.E., Manson J.E. and Gaziano J.M. Alcohol consumption and the risk of hypertension in women and men. Hypertension 2008;51:1080-1087. doi:10.1161/HYPERTENSIONAHA.107.104968
Writing Group of the PREMIER Collaborative Research Group. Effects of comprehensive lifestyle modification on blood pressure control: main results of the PREMIER Clinical Trial. JAMA 2003;289:2083-2093. doi:10.1001/jama.289.16.2083
World Health Organization, International Society of Hypertension Writing Group. 2003 World Health Organization (WHO)/International Society of Hypertension (ISH) statement on management of hypertension. Guidelines and recommendations. J Hyperten 2003;21:1983-1992.
Gli acidi grassi trans o grassi trans o TFA, acronimo dell’inglese trans fatty acids, sono acidi grassi insaturi, una sottoclasse di lipidi, con almeno un doppio legame in configurazione trans.
I doppi legami carbonio-carbonio hanno una conformazione planare e quindi possono essere considerati come dei piani ai cui lati opposti si attacca e prosegue la catena di atomi di carbonio. “L’ingresso” e “l’uscita” della catena può avvenire sullo stesso lato del piano, e in questo caso il doppio legame è detto in configurazione cis, o su lati opposti, e in quel caso si parla di configurazione trans. Questo è un esempio di isomeria geometrica o isomeria cis-trans.
Acidi Grassi trans, cis e saturi
Gli acidi grassi insaturi più comunemente hanno i loro doppi legami in configurazione cis; l’altra configurazione meno comune è appunto la trans.
Il legame cis determina una curvatura nella catena carboniosa dell’acido grasso, mentre la geometria del legame trans raddrizza la catena, impartendo alla molecola una struttura più simile a quella degli acidi grassi saturi.
Di seguito alcune caratteristiche distintive dei grassi a elevato tenore di acidi grassi trans, particolarmente vantaggiose per produrre margarine e grassi da pasticceria, ristorazione commerciale e per i processi di lavorazione industriale.
Le molecole curve non possono impacchettarsi facilmente mentre quelle lineari possono farlo. Ciò significa che i grassi trans concorrono, assieme ai geometricamente simili acidi grassi saturi, alla compattezza del grasso in cui si trovano conferendogli un punto di fusione più alto.
Innalzare il punto di fusione dei grassi significa che è possibile convertirli dalla forma liquida a quella semisolida o solida a temperatura ambiente.
Nota: gli acidi grassi trans tendono a essere meno solidi rispetto agli acidi grassi saturi.
Hanno, oltre a un punto di fusione, anche consistenza e gusto simili a quelli del burro.
Si conservano a lungo a temperatura ambiente.
Non vanno incontro a variazioni di sapore.
Sono stabili durante la frittura.
Origine
Gli acidi grassi trans presenti nella dieta possono essere di origine industriale, casalinga, vegetale o animale.
Nei paesi industrializzati la maggior parte dei grassi trans consumati nella dieta, negli Usa circa l’80% del totale, sono prodotti industrialmente, in quantità variabili, nel corso della parziale idrogenazione di oli alimentari ricchi di acidi grassi insaturi.
Sono prodotti a livello casalingo durante la frittura utilizzando oli vegetali contenenti acidi grassi insaturi.
Derivano dalla trasformazione batterica nel rumine dei ruminanti degli acidi grassi insaturi ingeriti dagli animali.
Un’altra fonte naturale è rappresentata da alcune piante come porri, piselli, lattuga e spinaci che contengono l’acido trans-3-esadecenoico, e l’olio di colza dove sono presenti l’acido brassidico (22:1∆13t) e l’acido gondoico (C20:1∆11t). In queste alimenti i grassi trans sono presenti in piccole quantità.
Piccolissime quantità, inferiori al 2%, si formano nel corso della deodorizzazione degli oli vegetali. Si tratta di un processo necessario nella raffinazione degli oli commestibili nel corso del quale si formano piccole quantità di acidi grassi trans con più di due doppi legami. Questi isomeri sono presenti anche nei cibi fritti e in quantità elevate in alcuni oli vegetali parzialmente idrogenati.
Acidi grassi trans industriali
L’idrogenazione è una reazione chimica nella quale l’idrogeno gassoso reagisce, in presenza di un catalizzatore, con una molecola.
L’idrogenazione degli acidi grassi insaturi consiste nell’addizione di atomi di idrogeno ai doppi legami presenti lungo la catena carboniosa degli acidi grassi. La reazione avviene in presenza di catalizzatori metallici ed è favorita dal riscaldamento degli oli vegetali in cui sono presenti i suddetti acidi grassi.
La parziale idrogenazione degli oli vegetali
Il processo di idrogenazione venne scoperto nel 1897 dal francese Paul Sabatier, premio Nobel per la chimica, insieme con il collega francese Victor Grignard, utilizzando come catalizzatore il nickel.
La parziale idrogenazione degli oli vegetali venne sviluppata nel 1903 da un chimico tedesco, Wilhelm Normann, che depositò un brevetto britannico intitolato “Process for converting unsaturated fatty acids or their glycerides into saturated compounds”. Il termine acidi grassi trans comparve per la prima volta nella Remark column della quinta edizione di “Standard Tables of Food Composition” in Giappone.
Nella parziale idrogenazione si verifica una saturazione incompleta dei doppi legami lungo la catena carboniosa dell’acido grasso insaturo. Ad esempio, per quanto riguarda l’olio di pesce il contenuto in acidi grassi trans in assenza di idrogenazione e in caso di forte idrogenazione è rispettivamente dello 0,5 e 3,6% mentre nel parziale idrogenazione è e del 30%.
Da Acido Oleico ad Acido Vaccenico
Ma, cosa più importante, alcuni dei doppi legami che restano possono cambiare posizione lungo la catena e produrre isomeri geometrici e di posizione, ossia i doppi legami possono essere modificati sia nella posizione che nella conformazione, con formazione appunto dei legami trans.
Inoltre si verificano anche le seguenti modificazioni.
Si formano monomeri ciclici e dimeri intramolecolari lineari.
Gli oli vegetali parzialmente idrogenati furono sviluppati per la produzione di grassi vegetali, un’alternativa più economica rispetto ai grassi animali. Infatti tramite questo processo, oli come quello di semi di lino, semi di cotone, germe di grano, soia, mais, girasole, sesamo, cartamo e vinacciolo, ricchi in acidi grassi insaturi, sono convertiti in grassi semisolidi.
Il primo olio idrogenato fu l’olio di semi di cotone negli USA nel 1911 per la produzione di grasso vegetale per pasticceria.
Negli anni ’30 la parziale idrogenazione divenne popolare con lo sviluppo della margarina.
Attualmente negli USA sono prodotte 2,7-3,6 milioni di tonnellate di oli idrogenati all’anno.
Acidi grassi trans dai ruminanti
Gli acidi grassi trans di origine animale sono prodotti dai batteri presenti nel rumine dei ruminanti, ad esempio mucche, pecore e capre, utilizzando come substrato una parte della relativamente piccola quantità degli acidi grassi insaturi contenuti negli alimenti di cui si nutrono, quindi mangimi, piante ed erbe.
E, considerando un animale che viva almeno un anno e abbia la possibilità di pascolare e/o mangiare fieno, esiste una variabilità stagionale che interessa gli acidi grassi insaturi assunti e i grassi trans prodotti. Infatti, in estate e primavera i pascoli possono contenere più acidi grassi insaturi rispetto ai mangimi che gli animali consumano durante l’inverno.
Si ritrovano di seguito in piccole ma significative quantità nei trigliceridi della carne e dei prodotti lattiero-caseari non magri, dove costituiscono tipicamente meno del 5% del totale degli acidi grassi. Nei trigliceridi, tali acidi grassi sono localizzati in posizione sn-1 e sn-3 della molecola, mentre nelle margarine e negli altri prodotti idrogenati di origine industriale sembrano essere concentrati in posizione sn-2.
Gli acidi grassi trans prodotti dai ruminanti sono principalmente acidi grassi monoinsaturi, con un numero di atomi di carbonio compreso tra 16 e 18, e costituiscono una piccola percentuale degli acidi grassi trans assunti con l’alimentazione (vedi sotto).
Isomeri
Sia per gli acidi grassi trans industriali che di origine animale, il gruppo più importante è rappresentato dagli isomeri trans del C18:1, ossia molecole contenenti 18 atomi di carbonio e un doppio legame trans la cui posizione varia tra il carbonio Δ6 e quello Δ16, con gli isomeri più comuni per entrambe le fonti con il doppio legame in posizione tra Δ9 e Δ11.
Tuttavia, sebbene una stessa molecola possa essere presente sia nel gruppo di derivazione industriale che animale, esiste una considerevole differenza quantitativa. Ad esempio, nel gruppo prodotto nei ruminanti, l’acido vaccenico (C18:1Δ11t) costituisce oltre il 60% degli isomeri trans-C18:1, mentre in quelli industriali l’acido elaidico (C18:1Δ9t) rappresenta il 15-20%, e il C18:1Δ10t e l’acido vaccenico oltre il 20% ciascuno.
Isomeri trans del C18:1
Effetti sulla salute
Gli acidi grassi trans che derivano dai ruminanti, nelle quantità effettivamente consumate nella dieta, non sono pericolosi per la salute umana.
Al contrario, il consumo dei grassi trans industriali non ha effetti benefici evidenti ne alcun valore intrinseco, oltre al loro contributo calorico, e, dal punto di vista della salute è solo dannoso, avendo effetti nocivi su:
livelli dei lipidi sierici;
cellule endoteliali;
processo infiammatorio;
altri fattori di rischio per le malattie cardiovascolari.
Inoltre sono associati positivamente con il rischio di cardiopatia coronarica, e di morte improvvisa per cause cardiache e diabete.
Nota: nel proseguo dell’articolo, se non diversamente specificato, ci si riferirà agli acidi grassi trans industriali, che verranno indicati semplicemente come acidi grassi trans o grassi trans.
Effetti a livello plasmatico
I livelli plasmatici del colesterolo LDL e del colesterolo HDL sono marcatori di rischio ben documentati per lo sviluppo di malattia coronarica o CHD, acronimo dell’inglese coronary heart disease:
elevati livelli di colesterolo LDL sono associati con un’aumentata incidenza di malattia cardiaca ischemica;
elevati livelli di colesterolo HDL si associano a una ridotta incidenza del rischio.
Per questo motivo il rapporto tra il livello del colesterolo totale e del colesterolo HDL è spesso usato come indicatore di rischio combinato per questi due componenti in relazione allo sviluppo della malattia cardiaca: maggiore è il rapporto, maggiore è il rischio.
I grassi trans hanno effetti nocivi sui lipidi sierici. Questi effetti sono stati valutati in numerosi studi di alimentazione controllata nei quali veniva effettuata una sostituzione isocalorica di acidi grassi saturi o cis-insaturi con grassi trans. E’ stato dimostrato che tale sostituzione:
aumenta livelli del colesterolo LDL;
riduce quelli del colesterolo HDL, a differenza degli acidi grassi saturi che invece aumentano i livelli di colesterolo HDL se utilizzati come sostituti in studi analoghi;
aumenta il rapporto tra colesterolo totale e colesterolo HDL, fino a valori pari a circa il doppio di quelli ottenuti con acidi grassi saturi, e, sulla base di questo solo effetto, è stato stimato che gli acidi grassi trans provochino circa il 6% degli eventi coronarici negli Stati Uniti.
Inoltre gli acidi grassi trans:
causano un pericoloso aumento delle LDL piccole e dense, che sono associate a un notevole aumento del rischio di CHD, anche in presenza di colesterolo LDL relativamente normale;
aumentano i livelli ematici dei trigliceridi, e questo è un fattore di rischio indipendente per la CHD;
aumentano i livelli della lipoproteina(a) o Lp(a), un altro importante fattore di rischio coronarico.
E tuttavia nel 2004 studi prospettici hanno dimostrato che la relazione tra l’assunzione di grassi trans e l’incidenza di CHD è maggiore di quella prevista da cambiamenti nei livelli dei lipidi sierici da soli. Ciò suggerisce che tali acido grassi influenzino altri fattori di rischio per la CHD, come l’infiammazione e la disfunzione delle cellule endoteliali.
Infiammazione e disfunzione delle cellule endoteliali
Il ruolo dell’infiammazione nell’aterosclerosi, e di conseguenza nella CHD, è cresciuto nell’ultimo decennio.
L’interleuchina-6, la proteina C-reattiva o CRP, acronimo dell’inglese C-reactive protein, e un’aumentata attività del fattore di necrosi tumorale o TNF, acronimo dell’inglese tumor necrosis factor, sono marcatori dell’infiammazione.
Nelle donne una maggiore assunzione di grassi trans è associata a un’aumentata attività del sistema TNF, e in quelle con un elevato indice di massa corporea con aumentati livelli di interleuchina-6 e CRP. Ad esempio, la differenza nei livelli di proteina C-reattiva osservata con un’assunzione media di grassi trans pari al 2,1% dell’assunzione di energia giornaliera totale, rispetto allo 0,9%, corrisponde a un aumento del rischio di malattia cardiovascolare del 30%. Risultati simili sono stati riportati in pazienti con malattia cardiaca, in studi randomizzati e controllati, in studi in vitro e in studi in cui sono stati analizzati i livelli di membrana degli acidi grassi trans, un biomarker della loro assunzione dietetica.
Quindi, i grassi trans promuovono l’infiammazione, e la loro azione pro-infiammatoria può spiegare parte dei loro effetti sulla CHD che, come visto, sono maggiori di quanto ci si aspetterebbe a seguito dei soli effetti sulle lipoproteine sieriche.
Attenzione: la presenza di infiammazione è un fattore di rischio indipendente non solo per la CHD ma anche per la resistenza all’insulina, il diabete, le dislipidemie e l’insufficienza cardiaca.
Un altro sito di azione degli acidi grassi trans può essere la funzione endoteliale. Diversi studi hanno suggerito l’associazione tra una loro maggiore assunzione e un aumento dei livelli dei biomarcatori circolanti della disfunzione endoteliale, come E-selectina, sICAM-1 e sVCAM-1.
Altri effetti
Studi in vitro hanno dimostrato che i grassi trans influenzano il metabolismo lipidico attraverso diversi percorsi.
Alterano la secrezione, la composizione lipidica e le dimensioni dell’apolipoproteina B-100 o Apo B-100.
Aumentano l’accumulo cellulare e la secrezione di colesterolo libero e di esteri del colesterolo da parte degli epatociti.
Alterano, negli adipociti, l’espressione dei geni per il PPAR-gamma, acronimo dell’inglese peroxisome proliferator-activated receptor gamma, la lipoprotein lipasi e la resistina, proteine con un ruolo centrale nel metabolismo degli acidi grassi e del glucosio.
Cardiopatia coronarica
Gli acidi grassi trans sono un fattore di rischio cardiovascolare indipendente.
Dai primi anni ’90 del secolo scorso l’attenzione si è focalizzata sui loro effetti sui lipidi plasmatici e sulla concentrazione delle lipoproteine.
Dal punto di vista epidemiologico, quattro importanti studi prospettici, che hanno coinvolto circa 140.000 soggetti monitorati per 6-14 anni, hanno tutti trovato riscontri positivi tra i livelli di questi acidi grassi nella dieta, valutati con l’aiuto di un questionario dettagliato sulla composizione della dieta, e il rischio di CHD.
I quattro studi sono:
The Health Professionals Follow-up study;
The Alpha-Tocopherol Beta-Carotene Cancer Prevention Study;
The Nurses’ Health Study;
The Zutphen Elderly Study.
Questi studi riguardano popolazioni talmente diverse che molto probabilmente i risultati sono applicabili all’intera popolazione mondiale.
Una meta-analisi degli studi suddetti ha mostrato che un 2% di aumento nell’apporto energetico dai grassi trans era associato con un incremento del 23% nell’incidenza di cardiopatia coronarica. Il rischio relativo di malattia cardiaca era di 1,36 nel “The Health Professionals Follow-up Study”, 1,14 nel “The Alpha-Tocopherol Beta-Carotene Cancer Prevention Study”; 1,93 (1,43-2,61) nel “The Nurses’ Health Study” e 1,28 (1,01-1,61) nel “The Zutphen Elderly Study”.
Quindi, anche un piccolo incremento del loro consumo si associa a un sostanziale aumento del rischio: il 2% dell’apporto energetico giornaliero, per una dieta di 2000 Kcal corrisponde a 40 Kcal o 4-5 grammi di grasso cioè l’equivalente di un cucchiaino di grasso!
Inoltre, in tre di questi studi l’associazione tra l’assunzione di grassi trans e il rischio di cardiopatia coronarica era più forte di una corrispondente associazione con l’assunzione di acidi grassi saturi. Nel “The Zutphen Elderly Study” questa associazione non è stata indagata.
A causa degli effetti negativi degli acidi grassi trans, per gli autori della meta-analisi non è etico condurre studi clinici randomizzati a lungo termine per saggiarne gli effetti sull’incidenza della cardiopatia coronarica. Per questi autori “studi osservazionali eseguiti in modo accurato forniscono un approccio ragionevole per valutarne gli effetti sulla salute cardiovascolare”.
Quindi, per evitare i loro effetti nocivi, di gran lunga peggiori in media a quelli dovuti ai contaminanti alimentari o ai residui dei pesticidi, è necessario evitarli o limitarne il consumo a meno dello 0,5% dell’apporto energetico giornaliero.
Ulteriori prove
Uno studio condotto su una popolazione australiana con un primo attacco di cuore e nessuna precedente storia di cardiopatia coronarica o iperlipidemia ha evidenziato un’associazione positiva tra i livelli di acidi grassi trans nel tessuto adiposo e il rischio di un infarto miocardico non fatale.
E’ stato dimostrato che la presenza nel tessuto adiposo di un acido grasso trans 18:1 con il doppio legame in posizione 7 (C18:1Δ7t), che si ritrova sia nei grassi animali che vegetali, era un predittore indipendente di un primo infarto miocardico, ossia il suo livello nel tessuto adiposo rimane un fattore predittivo per la malattia cardiaca dopo aggiustamento per i livelli di colesterolo totale. Di nuovo, sembra che solo una minima parte degli effetti negativi di grassi trans si attui per mezzo delle lipoproteine plasmatiche.
Nel corso di questo studio, a metà del 1996, i grassi trans furono eliminati dalle margarine vendute in Australia. Questa è stata un’opportunità unica per studiare la relazione temporale tra la loro l’assunzione e i loro livelli nel tessuto adiposo. E’ stato dimostrato che scomparivano dal tessuto adiposo sia dei pazienti che dei controlli con un tasso di circa il 15% del totale degli acidi grassi trans/anno.
Un altro studio condotto in Costarica ha trovato un’associazione positiva tra infarto miocardico e acidi grassi trans .
E’ interessante notare che in uno studio più grande, basato su una comunità caso-controllo, i livelli degli acidi grassi trans nelle membrane dei globuli rossi sono stati associati, dopo correzione per gli altri fattori di rischio, con un aumento del rischio di morte cardiaca improvvisa. Inoltre, l’aumento del rischio sembra essere correlato ai livelli dei trans-C18:2, i cui livelli sono stati associati con una triplicazione del rischio di morte improvvisa per cause cardiache, ma non con i livelli nella membrana cellulare di trans-C18:1, la principale classe di isomeri degli acidi grassi trans presenti nei cibi.
Diabete
In uno studio prospettico che ha coinvolto 84.204 infermiere di età compresa tra i 34 e i 59 anni, senza nessuna storia pregressa di diabete, malattia cardiovascolare e cancro, analizzate tra il 1980 e il 1996, e appartenenti al “The Nurses’ Health Study”, l’apporto di acidi grassi trans era correlato in modo significativo con il rischio di sviluppare il diabete di tipo 2. Dopo aggiustamenti per altri fattori di rischio l’apporto di grassi trans era positivamente associato con l’incidenza del diabete, con un incremento del rischio superiore al 39%.
Dati da studi di intervento controllati hanno mostrato che gli acidi grassi trans potrebbero danneggiare la sensibilità all’insulina in soggetti con resistenza all’insulina e diabete di tipo 2 più di quanto facciano gli acidi grassi insaturi, in particolare gli isomeri dell’acido linoleico coniugato o CLA, acronimo dell’inglese conjugated linoleic acid, trans-10,cis-12-CLA.
Attenzione perché alcuni integratori contengono isomeri del CLA e possono essere diabetogeni e proaterogenici in soggetti con resistenza all’insulina.
Nota: anche gli acidi grassi saturi innescano la medesima risposta, con nessuna differenza significativa tra loro e gli acidi grassi trans).
Nessun effetto significativo riguardo alla sensibilità all’insulina è stato invece osservato in soggetti sani e magri.
Acidi grassi trans dai ruminanti e cardiopatia coronarica
Quattro studi prospettici hanno valutato la relazione tra l’assunzione di acidi grassi trans dai ruminanti e il rischio di CHD: non è stata identificata alcuna associazione significativa.
In un altro studio pubblicato nel 2008 sono stati analizzati i dati provenienti da quattro studi di coorte danesi, che coinvolgevano 3686 adulti, arruolati tra il 1974 e il 1993, e seguiti per una mediana di 18 anni. In Danimarca il consumo di prodotti lattiero-caseari è relativamente alto e l’intervallo di assunzione dei grassi trans che provengono dai ruminati piuttosto ampio, sino al 1,1% dell’energia. Negli altri paesi, per la maggior parte delle persone, il consumo di grassi trans dai ruminati è sostanzialmente più basso del 1% dell’energia; negli USA è circa lo 0,5%.
Dopo correzione per altri fattori di rischio, non è stata trovata alcuna significativa associazione tra il loro consumo e l’incidenza di CHD, confermando, in una popolazione con un’assunzione relativamente alta di questo tipo di acidi grassi, le conclusioni dei quattro precedenti studi prospettici.
Quindi gli acidi grassi trans dai ruminanti, nelle quantità effettivamente consumate nella dieta, non aumentano il rischio di cardiopatia coronarica.
L’assenza di rischio rispetto ai grassi trans industriali può essere dovuta alla minore assunzione. Basti considerare che per gli Statunitensi la maggior parte degli acidi grassi trans sono di origine industriale, e che i livelli di grassi trans nel latte e nella carne sono relativamente bassi, dall’1% all’8% del totale dei grassi.
L’assenza di un rischio più elevato di CHD può essere dovuta anche alla presenza di isomeri diversi. Gli acidi grassi trans dai ruminanti e quelli industriali condividono molti isomeri, ma ci sono alcune differenze quantitative:
il livello di acido vaccenico è più alto nei grassi dei ruminanti, 30-50% degli isomeri trans-C18:1;
il gruppo del trans-C18:2, presente negli oli fritti, deodorinizzati e in alcuni oli parzialmente idrogenati, non lo è in quantità apprezzabile nel grasso dei ruminanti.
Infine altri fattori, ancora non noti e potenzialmente protettivi, potrebbero superare gli effetti nocivi dovuti ai grassi trans dai ruminanti.
Acidi grassi trans: esempi di legislazione in diversi paesi
USA Fino al 1985 non erano stati dimostrati effetti avversi dei grassi trans sulla salute umana e, addirittura, nel 1975 uno studio della Procter & Gamble non mostrò alcun effetto avverso sul colesterolo.
Il loro uso nei fast food crebbe dagli anni ’80, quando divenne chiaro il ruolo degli acidi grassi saturi nell’aumento del rischio cardiovascolare. Fu quindi condotta con successo una campagna per indurre la McDonald a passare dall’utilizzo del sego di bue a quello dell’olio vegetale per cucinare le patatine fritte. Nel frattempo diversi studi iniziarono a sollevare dubbi riguardo gli effetti dei grassi trans sulla salute.
La Food and Drug Administration, nel 1985, concluse che tali acidi grassi e l’acido oleico influenzavano il livello di colesterolo in maniera simile ma, dalla seconda metà del 1985 la pericolosità degli acidi grassi trans divenne chiara e la prova finale arrivò da studi di alimentazione controllata e da studi epidemiologici prospettici.
Nel 2003 la FDA ha stabilito che nei valori nutrizionali degli alimenti, sia per i cibi classici che per gli integratori, sia mostrato il loro contenuto a partire dal primo gennaio 2006. In particolare, questa ordinanza rappresenta il primo sostanziale cambiamento alla etichettatura dei prodotti alimentari dall’obbligo, risalente al 1990, di mettere le informazioni per porzione.
Nel 2005 il Dipartimento dell’Agricoltura statunitense ha fatto di un apporto minimo di grassi trans una raccomandazione chiave della nuova piramide alimentare.
Nel 2006 l’American Heart Association raccomanda di limitarne l’apporto all’1% del consumo calorico giornaliero e suggerisce ai produttori di alimenti e ai ristoranti di passare ad altri grassi.
Sempre nel 2006, il Dipartimento della Sanità della città di New York annuncia il divieto del loro utilizzo nei suoi 40.000 ristoranti entro il primo giugno 2008, seguito nel 2010-2011 dallo stato della California.
Australia Dopo il giugno 1996 sono stati eliminati dalla margarina venduta in Australia, che in passato contribuiva per circa il 50% al loro apporto giornaliero in quel paese.
Europa
Dall’11 marzo 2003 il governo danese, dopo un dibattito iniziato nel 1994 e due nuovi rapporti nel 2001 e nel 2003, ha deciso di eliminarne gradualmente l’uso nei cibi prima della fine del 2003.
Due anni più tardi però, la Commissione Europea ha chiesto alla Danimarca di ritirare questa legge che, sfortunatamente, non è stata accettata a livello comunitario.
Tuttavia, nel 2007, la stessa Commissione Europea ha ritirato la procedura d’infrazione nei confronti della Danimarca grazie alle maggiori prove scientifiche sulla pericolosità di questo tipo di acidi grassi.
L’esempio danese è stato seguito nel 2009 dall’Austria e dalla Svizzera, nel 2011 dall’Islanda, dalla Norvegia e Ungheria nel 2014, e più di recente Estonia e Georgia. Quindi circa il 10% della popolazione della Comunità Europea, circa 500 milioni di persone, vive in paesi dove è illegale vendere prodotti con un elevato contenuto di grassi trans. I governi degli altri paesi della Comunità Europea fanno invece affidamento sulla volontà dei produttori di alimenti di ridurne il contenuto nei propri prodotti. Tale strategia si è dimostrata efficace solo nei paesi dell’Europa Occidentale.
Canada
Il Canada sta considerando la legislazione per eliminarli dalle forniture alimentari e nel 2005 ha introdotto l’obbligo di mostrare il loro contenuto negli alimenti preconfezionati.
In conclusione, con l’eccezione di pochi paesi nei quali l’uso a scopi alimentari dei grassi trans è stato vietato, negli altri l’unico modo per ridurne l’apporto è la scelta del consumatore di preferire cibi in cui non siano presenti, leggendo sempre gli ingredienti, anche nel caso in cui il loro contenuto sulla confezione sia dichiarato pari a 0, potendo provenire dalla margarina, dai grassi vegetali per pasticceria, dall’olio vegetale e dalla frittura. Infatti ad esempio negli USA, i produttori di alimenti che ne contengano meno di 0,5 g per porzione possono dichiararne sulla confezione un contenuto pari a 0. Questo contenuto è basso, ma se un consumatore mangia più porzioni, ne consuma quantità sostanziali.
Attenzione: la lista degli ingredienti non è obbligatoria nei ristoranti, forni e in molti altri esercizi alimentari al dettaglio.
Riformulazione degli alimenti per ridurre gli acidi grassi trans
Le organizzazioni di sanità pubblica, compresa l’Organizzazione Mondiale della Sanità a partire dal settembre del 2006, hanno raccomandato di ridurre il consumo di acidi grassi trans. Solo negli USA la loro quasi eliminazione potrebbe evitare tra i 72.000 e i 280.000 casi di malattie cardiovascolari degli 1,2 milioni che si verificano annualmente.
I produttori di alimenti e i ristoranti possono ridurre l’uso dei grassi trans scegliendo alternative agli oli parzialmente idrogenati.
In Danimarca la loro eliminazione dagli oli vegetali non ha aumentato il consumo dei grassi saturi in quanto sono stati per lo più sostituiti con acidi grassi cis-insaturi. Inoltre, non ci sono stati effetti evidenti per i consumatori: ne aumenti nei costi ne riduzioni della disponibilità e della qualità dei prodotti alimentari.
Nel 2009, Stender e colleghi, analizzando ciò che è accaduto negli USA, in Sud Africa e molte nazioni europee, hanno dimostrato che gli acidi grassi trans in alimenti come patatine fritte, biscotti, torte e pop corn per il microonde possono essere sostituiti, a un prezzo simile, con una miscela di acidi grassi saturi, monoinsaturi e polinsaturi. Una tale sostituzione assicura anche maggiori benefici nutrizionali rispetto a quella uno a uno tra i grassi trans e i grassi saturi da soli.
Tuttavia attenzione perché ad esempio solamente nelle patatine fritte con bassi livelli di grassi trans la percentuale di acidi grassi saturi rimane costante, mentre nei biscotti e nelle torte è in media del 33% maggiore e nei pop corn per forno a microonde del 24%: gli acidi grassi saturi sono si meno pericolosi degli grassi trans ma di più rispetto agli acidi grassi mono- e polinsaturi.
Lo stesso gruppo di ricerca, analizzando alcuni cibi di comune consumo in Europa, acquistati in supermercati, anche della stessa catena, e in fast food (McDonald’s e Kentucky Fried Chicken, KFC) dal 2005 al 2014, ha dimostrato come il loro contenuto in grassi trans sia ridotto o addirittura assente in diversi paesi dell’Europa Occidentale mentre rimanga ancora alto in paesi dell’Europa dell’Est e del Sud-Est.
Nel 2010 Mozaffarian e colleghi hanno valutato i livelli degli acidi grassi trans e dei grassi saturi nei cibi dei principali supermercati di marca statunitensi e dei ristoranti dopo la riformulazione per ridurre il contenuto dei primi in due momenti: dal 1993 sino al 2006 e dal 2006 al 2009. E’ stata rilevata una generale riduzione del loro contenuto senza alcun sostanziale od equivalente aumento nel contenuto degli acidi grassi saturi.
Fonti alimentari
Nel mondo sono molti gli alimenti ricchi di grassi trans comunemente consumati.
Negli USA gran parte proviene da oli vegetali parzialmente idrogenati, con un consumo medio da questa fonte che è rimasto costante dagli anni ’60 dello scorso secolo.
Da notare che i valori degli acidi grassi trans che si incontreranno devono essere interpretati con cautela perché molti fast food, ristoranti e industrie possono aver cambiato il tipo di grasso utilizzato per la frittura e la cottura da quando gli articoli analizzati sono stati pubblicati.
I valori riportati, se non diversamente specificato, si riferiscono al contenuto percentuale in acidi grassi trans/100 g di acidi grassi.
Margarina
Tra gli alimenti ricchi di grassi trans la margarina, in bastoncini o dura, ne ha avuto la percentuale più alta. Tuttavia, i loro livelli hanno iniziato a ridursi quando il miglioramento della tecnologia ha permesso la produzione di margarine più morbide, che nel tempo sono divenute popolari. Esistono comunque differenze nel contenuto di grassi trans di margarine provenienti da diversi paesi. Di seguito alcuni esempi.
Il contenuto più alto, 13-16,5%, si ritrova nelle margarine morbide provenienti dall’Islanda, Norvegia e Regno Unito.
Un contenuto minore è presente in quelle dall’Italia, Germania, Finlandia e Grecia, rispettivamente il 5,1%, 4,8%, 3,2%, e 2,9%.
In Portogallo, Paesi Bassi, Belgio, Danimarca, Francia, Spagna e Svezia il contenuto è inferiore al 2%.
USA e Canada sono in ritardo rispetto all’Europa, ma negli USA sono in corso dei cambiamenti a seguito dell’introduzione dei grassi trans nell’etichettatura degli alimenti e della maggior conoscenza dei pericoli associati al loro consumo da parte degli acquirenti. Per questo motivo, al momento, negli USA la margarina è considerata contribuire in misura minore al totale dei grassi trans, mentre le fonti principali sono i prodotti commerciali da forno e dei fast food, come torte, biscotti, wafer, cracker, crocchette di pollo, patatine fritte o pop corn per il forno a microonde (vedi sotto).
Grassi da pasticceria
Il contenuto in grassi trans dei grassi da pasticceria è compreso tra il 6% e il 50%, e varia nei diversi paesi: in Germania, Austria e Nuova Zelanda è più basso che in Francia o negli USA.
Tuttavia, come per la margarina, anche per i grassi da pasticceria il loro contenuto è in diminuzione. In Germania è calato dal 12% nel 1994 al 6% nel 1999, in Danimarca è il 7% (1996), mentre in Nuova Zelanda è di circa il 6% (1997).
Oli vegetali
Al momento gli oli vegetali non idrogenati utilizzati per condire le insalate o per cucinare non contengono acidi grassi trans o ne contengono solo piccole quantità.
La lavorazione di questi oli può comportarne la produzione di piccole quantità, variabili da 0,05 g/100 g di prodotto per l’olio di oliva extra-vergine a 2,42 g/100 g di prodotto per l’olio di colza. Quindi il loro contributo al contenuto di grassi trans dei cibi attualmente consumati è molto piccolo.
Un’eccezione è rappresentata dagli oli vegetali idrogenati pachistani, che ne possono contenere dal 14% al 34%.
Minestre pronte
Tra gli alimenti ricchi di grassi trans le minestre pronte ne contengono quantità significative, che variano dal 10% del brodo di manzo al 35% della crema di cipolle; quindi, se consumate di frequente contribuiscono in grande misura all’apporto di tali acidi grassi nella dieta.
Prodotti alimentari trasformati
Gli acidi grassi trans, grazie alle loro proprietà sono utilizzati nella preparazione di molti alimenti trasformati come biscotti, torte, cornetti, dolci e altri prodotti da forno. E, nella dieta consumata in Nord America, i prodotti da forno ne sono la principale fonte.
Naturalmente, il contenuto in grassi trans dipende dal tipo di grassi utilizzati nella lavorazione.
Salse
La maionese, i condimenti per le insalate e altre salse apportano quantità di grassi trans piccole o nulle.
Latte umano e alimenti per neonati
Il contenuto di acidi grassi trans del latte umano riflette il contenuto in tali grassi della dieta materna nel giorno precedente, è compreso tra 1% e il 7% ed è in diminuzione dal 7,1% del 1998 al 4,6% del biennio 2005-2006.
Gli alimenti per lattanti hanno valori di grassi trans in media di 0,1%-4,5%, con una marca che raggiunge il 15,7%.
Gli alimenti per bambini ne contengono più del 5%.
Fast food e ristoranti
Nei fast food e ristoranti, per la frittura, vengono utilizzati grassi usati anche in pasticceria, quindi con elevate quantità di acidi grassi trans; pertanto i cibi venduti possono contenerne quantità relativamente alte. Fonti sono le torte fritte, le patatine fritte, le crocchette di pollo, gli hamburger, le fritture di pesce e il pollo fritto.
In lavori pubblicati da Stender e colleghi dal 2006 al 2009 viene dimostrato che il contenuto di grassi trans delle patatine fritte e delle crocchette di pollo variava ampiamente da paese a paese ma, anche all’interno della stessa catena di fast food, in uno stesso paese, e addirittura in una stessa città, a causa dell’olio utilizzato per cucinare. Ad esempio, l’olio utilizzato per le patatine fritte nei punti vendita di McDonald’s negli USA e in Perù ne conteneva il 23-24%, mentre l’olio utilizzato nella stessa catena in molti paesi europei ne contiene circa il 10%, con alcuni paesi, come la Danimarca, a un livello più basso del 5% e 1%.
E, considerando un pasto a base di patatine fritte e crocchette di pollo, in porzioni rispettivamente da 171 e 160 g, acquistato da McDonald a New York, lo stesso conteneva oltre 10 g di grassi trans mentre, se veniva acquistato da KFC in Ungheria si arrivava a quasi a 25 g.
Di seguito, sempre dai lavori di Stender e colleghi, si può osservare una comparazione tra diversi paesi per quanto riguarda il contenuto di grassi trans delle crocchette di pollo e delle patatine fritte acquistate da McDonald o da KFC.
Crocchette di pollo e patatine fritte da McDonald:
meno di un grammo solo se i pasti sono stati acquistati in Danimarca;
1-5 g in Portogallo, Paesi Bassi, Russia, Repubblica Ceca o Spagna;
5-10 g negli USA, Perù, Regno Unito, Sud Africa, Polonia, Finlandia, Francia, Italia, Norvegia, Spagna, Svezia, Germania o Ungheria.
Crocchette di pollo e patatine fritte da KFC:
meno di 2 g se i pasti sono stati acquistati nel Regno Unito, Danimarca, Russia o Germania;
2-5 g in Germania, Francia, Regno unito, Spagna o Portogallo;
5-10 g alle Bahamas, Sud Africa o USA;
10-25 g in Ungheria, Polonia, Perù o Repubblica Ceca.
Bibliografia
Akoh C.C. and Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 3rd Edition. CRC Press Taylor & Francis Group, 2008
Ascherio A., Katan M.B., Zock P.L., Stampfer M.J., Willett W.C. Trans fatty acids and coronary heart disease. N Engl J Med 1999;340:1994-1998. doi:10.1056/NEJM199906243402511
Ascherio A., Rimm E.B., Giovannucci E.L., Spiegelman D., Stampfer M., Willett W.C. Dietary fat and risk of coronary heart disease in men: cohort follow up study in the United States. BMJ 1996; 313:84-90. doi:10.1080/17482970601069094
Asp N-G. Fatty acids in focus – the good and the bad ones. Scand J Food Nutr 2006;50:155-160. doi:10.1080/17482970601069094
Baylin A., Kabagambe E.K., Ascherio A., Spiegelman D., Campos H. High 18:2 trans-fatty acids in adipose tissue are associated with increased risk of nonfatal acute myocardial infarction in Costa Rican adults. J Nutr 2003;133:1186-1191. doi:10.1093/jn/133.4.1186
Chow C.K. Fatty acids in foods and their health implication. 3rd Edition. CRC Press Taylor & Francis Group, 2008.
Clifton P.M., Keogh J.B., Noakes M. Trans fatty acids in adipose tissue and the food supply are associated with myocardial infarction. J Nutr 2004;134:874-879. doi:10.1093/jn/134.4.874
Costa N., Cruz R., Graça P., Breda J., and Casal S. Trans fatty acids in the Portuguese food market. Food Control 2016;64:128-134. doi:10.1016/j.foodcont.2015.12.010
Eckel R.H., Borra S., Lichtenstein A.H., Yin-Piazza D.Y. Understanding the Complexity of Trans fatty acid reduction in the American diet. American Heart Association trans fat conference 2006 report of the trans fat conference planning group. Circulation 2007;115:2231-2246. doi:10.1161/CIRCULATIONAHA.106.181947
Hu F.B., Manson J.E., Stampfer M.J., Colditz G., Liu S., Solomon C.G., and Willett W.C. Diet, lifestyle, and the risk of type 2 diabetes mellitus in women. N Engl J Med 2001;345:790-797. doi:10.1056/NEJMoa010492
Hu F.B., Willett W.C. Optimal diet for prevention of coronary heart disease JAMA 2002;288:2569-2578. doi:10.1001/jama.288.20.2569
Lemaitre R.N., King I.B., Raghunathan T.E., Pearce R.M., Weinmann S., Knopp R.H., Copass M.K., Cobb L.A., Siscovick D.S. Cell membrane trans-fatty acids and the risk of primary cardiac arrest. Circulation 2002;105:697-701. doi:10.1161/hc0602.103583
Lemaitre R.N, King I.B, Mozaffarian D., Sootodehnia N., Siscovick D.S. Trans-fatty acids and sudden cardiac death. Atheroscler Suppl 2006; 7(2):13-5. doi:10.1016/j.atherosclerosissup.2006.04.003
Lichtenstein A.H. Dietary fat, carbohydrate, and protein: effects on plasma lipoprotein patterns J. Lipid Res. 2006;47:1661-1667. doi:10.1194/jlr.R600019-JLR200
Lichtenstein A.H., Ausman L., Jalbert S.M., Schaefer E.J. Effect of different forms of dietary hydrogenated fats on serum lipoprotein cholesterol levels. N Engl J Med 1999;340:1933-1940. doi:10.1056/NEJM199906243402501
Lopez-Garcia E., Schulze M.B., Meigs J.B., Manson J.E, Rifai N., Stampfer M.J., Willett W.C. and Hu F.B. Consumption of trans fatty acids is related to plasma biomarkers of inflammation and endothelial dysfunction. J Nutr 2005;135:562-566. doi:10.1093/jn/135.3.562
Masanori S. Trans Fatty Acids: Properties, Benefits and Risks J Health Sci 2002;48(1):7-13.
Mensink R.P., Katan M.B. Effect of dietary trans fatty acids on high-density and low-density lipoprotein cholesterol levels in healthy subjects. N Engl J Med 1990;323:439-445. doi:10.1056/NEJM199008163230703
Mozaffarian D. Commentary: Ruminant trans fatty acids and coronary heart disease-cause for concern? Int J Epidemiol 2008;37(1):182-184. doi:10.1093/ije/dym263
Mozaffarian D., Jacobson M.F., Greenstein J.S. Food Reformulations to reduce trans fatty acids. N Eng J Med 2010;362:2037-2039. doi:https://doi.org/10.1056/NEJMc1001841
Mozaffarian D., Katan M.B., Ascherio A., Stampfer M.J., Willett W.C. Trans fatty acids and cardiovascular disease. N Engl J Med 2006;354:1601-1613. doi:10.1056/NEJMra054035
Mozaffarian D., Pischon T., Hankinson S.E., Joshipura K., Willett W.C., and Rimm E.B. Dietary intake of trans fatty acids and systemic inflammation in women. Am J Clin Nutr 2004;79:606-612. doi:https://doi.org/10.1093/ajcn/79.4.606
Oh K., Hu F.B., Manson J.E., Stampfer M.J., Willett W.C. Dietary fat intake and risk of coronary heart disease in women: 20 years of follow-up of the Nurses’ Health Study. Am J Epidemiol 2005;161(7):672-679. doi:10.1093/aje/kwi085
Okie S. New York to Trans Fats: You’re Out! N Engl J Med 2007;356:2017-2021. doi:10.1056/NEJMp078058
Oomen C.M., Ocke M.C., Feskens E.J., van Erp-Baart M.A., Kok F.J., Kromhout D. Association between trans fatty acid intake and 10-year risk of coronary heart disease in the Zutphen Elderly Study: a prospective population-based study. Lancet 2001; 357(9258):746-751. doi:10.1016/S0140-6736(00)04166-0
Pietinen P., Ascherio A., Korhonen P., Hartman A.M., Willett W.C., Albanes D., VirtamO J.. Intake of fatty acids and risk of coronary heart disease in a cohort of Finnish men: the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study. Am J Epidemiol 1997;145(10):876-887. doi:10.1093/oxfordjournals.aje.a009047
Risérus U. Trans fatty acids, insulin sensitivity and type 2 diabetes. Scand J Food Nutr 2006;50(4):161-165. doi:10.1080/17482970601133114
Salmerón J., Hu F.B., Manson J.E., Stampfer M.J., Colditz G.A., Rimm E.B., and Willett W.C. Dietary fat intake and risk of type 2 diabetes in women. Am J Clin Nutr 2001;73:1019-1026 doi:10.1093/ajcn/73.6.1019
Stender S., Astrup A., and Dyerberg J. A trans European Union difference in the decline in trans fatty acids in popular foods: a market basket investigation. BMJ Open 2012;2(5):e000859. doi:10.1136/bmjopen-2012-000859
Stender S., Astrup A., and Dyerberg J. Artificial trans fat in popular foods in 2012 and in 2014: a market basket investigation in six European countries. BMJ Open 2016;6(3):e010673. doi:10.1136/bmjopen-2015-010673
Stender S., Astrup A.,and Dyerberg J. Tracing artificial trans fat in popular foods in Europe: a market basket investigation. BMJ Open 2014;4(5):e005218. doi:10.1136/bmjopen-2014-005218
Stender S., Astrup A., Dyerberg J. What went in when trans went out?. N Engl J Med 2009;361:314-316. doi:10.1056/NEJMc0903380
Stender S., Dyerberg J. The influence of trans fatty acids on health. Fourth edition. The Danish Nutrition Council; publ. no. 34, 2003.
Stender S., Dyerberg J., Astrup A. Consumer protection through a legislative ban on industrially produced trans fatty acids in Denmark. Scand J Food Nutr 2006;50(4):155-160. doi:10.1080/17482970601069458
Stender S., Dyerberg J., Astrup A. High levels of trans fat in popular fast foods. N Engl J Med 2006;354:1650-1652. doi:10.1056/NEJMc052959
Willett W., Mozaffarian D. Ruminant or industrial sources of trans fatty acids: public health issue or food label skirmish? Am J Clin Nutr 2008;87(3): 515-516. doi:10.1093/ajcn/87.3.515
L’assunzione di carboidrati può migliorare la capacità di resistenza e la prestazione.
L’ingestione di diversi tipi di carboidrati, che utilizzano trasportatori intestinali differenti, può:
aumentare l’assorbimento totale dei carboidrati;
aumentare l’ossidazione dei carboidrati assunti;
migliorare la prestazione.
Dagli studi condotti la migliore combinazione è risultata essere maltodestrine e fruttosio nel rapporto 2:1.
Glucosio e fruttosio
Quando durante l’esercizio fisico prolungato viene assunta una miscela di glucosio e fruttosio, nella letteratura analizzata rispettivamente 1,2 e 0,6 g/min, rapporto 2:1, per una velocità di assunzione complessiva pari a 1,8 g/min, c’è una minor competizione per l’assorbimento intestinale rispetto all’ingestione di una quantità isoenergetica di solo glucosio o solo fruttosio, essendo coinvolti due trasportatori intestinali differenti. Inoltre, l’assorbimento del fruttosio è stimolato dalla presenza del glucosio.
Tutto ciò può:
aumentare la disponibilità di carboidrati esogeni nel sangue;
produrre una velocità di ossidazione dei carboidrati esogeni maggiore rispetto al solo glucosio.
Dalla coingestione di glucosio e fruttosio si ottiene una velocità di ossidazione dei carboidrati esogeni di circa 1,26 g/min, quindi maggiore rispetto a quella osservata con l’assunzione del solo glucosio (1 g/min) anche in alte concentrazioni.
La differenza osservata, ossia +0,26 g/min, può essere attribuita per intero all’ossidazione del fruttosio ingerito.
Saccarosio e glucosio
L’ingestione di saccarosio e glucosio, nelle stesse condizioni dell’ingestione di glucosio e fruttosio, quindi rispettivamente 1,2 e 0,6 g/min, in rapporto 2:1, per apporto complessivo di carboidrati pari a 1,8 g/min, dà risultati simili.
Glucosio, saccarosio e fruttosio
Con la combinazione di glucosio, saccarosio e fruttosio si ottengono velocità di ossidazione molto elevate (nella letteratura analizzata rispettivamente 1,2, 0,6 e 0,6 g/min, in rapporto 2:1:1, per apporto complessivo di carboidrati pari a 2,4 g/min; tuttavia, notare la maggiore quantità di carboidrati assunta).
Maltodestrine e fruttosio
Velocità di ossidazione elevate si osservano anche con combinazioni di maltodestrine e fruttosio, nelle stesse condizioni dell’ingestione di glucosio e fruttosio (quindi rispettivamente 1,2 e 0,6 g/min, in rapporto 2:1, per apporto complessivo di carboidrati pari a 1,8 g/min).
Queste elevate velocità di ossidazione possono essere raggiunte con carboidrati disciolti in una bevanda, presenti in un gel o in barrette a basso contenuto di grasso, proteina e fibra.
La migliore combinazione di carboidrati da assumere durante l’esercizio fisico prolungato è probabilmente la miscela di maltodestrine e fruttosio in rapporto 2:1, in una soluzione al 5%, per un apporto di circa 80-90 g/h.
Ossidazione dei Carboidrati Ingeriti
Perche?
Questa miscela ha il miglior rapporto tra la quantità di carboidrati ingerita e la loro velocità di ossidazione e questo significa che quantità più piccole di carboidrati rimangono nello stomaco o nell’intestino riducendo il rischio di complicanze/disturbi gastrointestinali durante esercizio prolungato.
Una soluzione che contenga diversi tipi di carboidrati e che ne abbia un contenuto non superiore al 5% ottimizza lo svuotamento gastrico e migliora l’apporto di liquidi.
Esempi di soluzioni di carboidrati al 5% contenenti circa 80-90 g di maltodestrine e fruttosio in rapporto 2:1; tempo di ingestione di circa un’ora:
1,5 L di soluzione: 80 g di carboidrati, rispettivamente circa 55 g di maltodestrine e circa 25 g di fruttosio.
1,8 L di soluzione: 90 g di carboidrati, rispettivamente 60 g di maltodestrine e 30 g di fruttosio.
Conclusioni
Durante l’esercizio fisico prolungato, quando sono necessarie elevate velocità di ossidazione dei carboidrati esogeni, è preferibile l’ingestione di carboidrati differenti rispetto a quella di grandi quantità di un singolo carboidrato.
La migliore combinazione sembra essere quella tra maltodestrine e fruttosio, in rapporto di 2:1, in una soluzione al 5%, e con una velocità di ingestione di circa 80-90 g/h.
Bibliografia
Burke L.M., Hawley J.A., Wong S.H.S., & Jeukendrup A. Carbohydrates for training and competition. J Sport Sci 2011;29:Sup1,S17-S27. doi:10.1080/02640414.2011.585473
Jentjens R.L.P.G., Moseley L., Waring R.H., Harding L.K., and Jeukendrup A.E. Oxidation of combined ingestion of glucose and fructose during exercise. J Appl Physiol 2004:96;1277-1284. doi:10.1152/japplphysiol.00974.2003
Jentjens R.L.P.G., Venables M.C., and Jeukendrup A.E. Oxidation of exogenous glucose, sucrose, and maltose during prolonged cycling exercise. J Appl Physiol 2004:96;1285-1291. doi:10.1152/japplphysiol.01023.2003
Jeukendrup A.E. Carbohydrate feeding during exercise. Eur J Sport Sci 2008:2;77-86. doi:10.1080/17461390801918971
Jeukendrup A.E. Nutrition for endurance sports: marathon, triathlon, and road cycling. J Sport Sci 2011:29;sup1, S91-S99. doi:10.1080/02640414.2011.610348
I carotenoidi sono una classe di pigmenti liposolubili ampiamente presenti in natura.
Sono composti organici di colore giallo, arancio e rosso, sono costituiti da 8 unità isopreniche e hanno numerosi doppi legami coniugati. La loro catena idrocarburica può andare incontro a modificazioni che ne influenzano significativamente le proprietà biologiche.
Scoperti nella prima metà del 1800, i carotenoidi sono prodotti da tutti gli organismi fotosintetici, comprese piante, macroalghe e microalghe. Inoltre sono sintetizzati anche da organismi non fotosintetici quali alcuni tipi di funghi, batteri e insetti come gli Afidi dei piselli, il ragnetto rosso, una specie di acaro, e alcune specie di moscerini.
Con polifenoli e glucosinolati, costituiscono il gruppo dei fitochimici.
Nel corso dell’evoluzione, grazie alle loro proprietà chimiche e fisiche, si sono dimostrati estremamente versatili, essendo in grado di svolgere molte funzioni sia nelle piante che negli animali, tra le quale è molto importante l’azione antiossidante. Inoltre, nell’uomo molti carotenoidi sono precursori della vitamina A.
Si contano più di 750 carotenoidi differenti, più di 100 sono stati trovati nella frutta e verdura, e circa 40 sono assunti in quantità significative dall’uomo.
Grazie al loro colore, i carotenoidi sono utilizzati come additivi alimentari.
I carotenoidi sono stati scoperti nella prima metà del diciannovesimo secolo.
Il primo carotenoide a essere isolato fu il beta-carotene nel 1831, grazie al lavoro del farmacista tedesco Heinrich Wilhelm Ferdinand Wackenroder, il quale chiamò carotene il pigmento giallo che riuscì a cristallizzare dalla radice della carota.
Jöns Jacob Berzelius, considerato uno dei padri della chimica moderna, chiamò xantofille, dal greco xanthos, che significa giallo, e phyllon che significa foglia, i pigmenti gialli estratti dalle foglie autunnali.
Mikhail Semyonovich Tsvet , botanico italo-russo, che nel 1906 ideò la cromatografia, riuscì a separare i pigmenti fogliari, ossia le clorofille dai caroteni e xantofille, grazie alla cromatografia su colonna, e chiamò carotenoidi i pigmenti dal giallo all’arancio.
Struttura chimica
I carotenoidi sono una classe di lipidi aventi come unità strutturale di base una sequenza lineare di otto unità isopreniche. Sono quindi terpeni, precisamente tetraterpeni, per un totale di 40 atomi di carbonio.
Le unità isopreniche sono unite con un relazione posizionale 1,5, ossia testa-coda, tranne al centro della catena idrocarburica, dove la relazione posizionale è 1,6, quindi coda-coda, il che, invertendo l’ordine, rende la molecola simmetrica.
I carotenoidi sono caratterizzati dalla presenza di un esteso sistema di doppi legami, dei quali da 3 a 13 possono essere coniugati.
La catena idrocarburica dei carotenoidi può andare incontro a modificazioni chimiche quali idrogenazione, deidrogenazione, introduzione di atomi di ossigeno, ciclizzazione di una o entrambe le estremità a formare anelli ionone, ed esterificazione con acidi grassi. Tali modificazioni permettono la formazione di molte strutture differenti, la maggior parte delle quali sono descritte dalla formula chimica C40H56On, con 0≤n≤6.
Tradizionalmente, ai carotenoidi sono stati assegnati nomi comuni provenienti dalla fonte biologica da cui sono stati isolati, sebbene possano essere denominati anche utilizzando la nomenclatura IUPAC.
Isomeria
I carotenoidi presentano molti doppi legami che possono avere isomeriacis–trans, anche detta isomeria geometrica. Sebbene ognuno dei doppi legami possa essere in configurazione cis o trans, in natura la configurazione prevalente è la trans, termodinamicamente più stabile rispetto alla configurazione cis grazie al minor ingombro sterico tra i sostituenti.
La rotazione intorno ai legami semplici, o legami sigma, presenti lungo la catena carboniosa permette anche la formazione di isomeri conformazionali. Quando i doppi legami sono presenti sullo stesso lato rispetto al legame sigma si parla di conformazione s-cis. Al contrario, se i doppi legami sono su lati opposti rispetto al legame sigma si parla di conformazione s-trans.
In natura i carotenoidi aciclici sono principalmente in conformazione s-trans, in quanto è quella a minor ingombro sterico. In presenza di strutture cicliche alle estremità della catena del carotenoide, la rotazione intorno al legame sigma C6-C7 da luogo alla formazione di un isomero 6-s-cis a minor ingombro sterico e quindi energeticamente favorito.
Si noti che l’ingombro sterico e le dimensioni dei carotenoidi influenzano la capacità di interagire con enzimi o di formare strutture sovramolecolari.
La maggior parte dei carotenoidi presenta anche un centro di chiralità o un asse di chiralità, che fa si che esistano isomeri ottici. In questi casi, per dare un nome univoco alla molecola si ricorre al sistema RS, e non alle regole della convenzione di Fischer, utilizzata solo per carboidrati e aminoacidi.
Classificazione
I carotenoidi possono essere classificati sulla base della presenza o meno di ossigeno o di strutture cicliche.
Sulla base della presenza o meno di ossigeno sono suddivisi in xantofille e caroteni.
I caroteni, privi di ossigeno, sono formati da soli atomi di carbonio e idrogeno, e hanno formula chimica C40H56. Esempi di caroteni sono l’alfa-carotene, il beta-carotene, il delta-carotene, lo zeta-carotene, il fitoene, e il licopene.
Le xantofille contengono anche atomi di ossigeno, hanno formula chimica C40H56On, con 0˂n≤6, e possono a loro volta essere suddivise in:
idrossicarotenoidi, che contengono almeno un gruppo idrossilico, come l’alfa-criptoxantina, la beta-criptoxantina, la zeaxantina e la luteina;
epossicarotenoidi, che presentano almeno un gruppo epossidico, come l’anteraxantina, l’auroxantina e la luteoxantina;
chetocarotenoidi, che presentano uno o più gruppi carbonilici, come l’astaxantina e la cantaxantina.
La presenza o meno di strutture cicliche nella molecola permette la suddivisione in carotenoidi ciclici e aciclici.
I carotenoidi ciclici contengono una o due strutture cicliche, e, rispetto agli aciclici sono più corti, ma con un ingombro sterico maggiore e un maggior spazio occupato. Esempi sono l’alfa-carotene, il beta-carotene, il gamma-carotene e il delta-carotene.
I carotenoidi aciclici sono molecole formate da una catena carboniosa lineare. Esempi sono il licopene, lo zeta-carotene, il fitoene, e il fitofluene.
Infine esistono anche carotenoidi non comuni o specie-specifici, come la bixina, la capsantina e la capsorubina.
Solubilità
I carotenoidi sono molecole idrofobiche, quindi solubili in solventi organici, solubilità che varia in funzione dei sostituenti presenti. Data lo loro idrofobicità, nelle cellule si ritrovano nelle membrane cellulari. Nella maggior parte dei casi le xantofille presentano i gruppi polari ai lati opposti della catena, e, quando presenti nelle membrane cellulari, al fine di minimizzare l’energia del sistema, si dispongono in modo tale che i gruppi polari siano in contato con i gruppi polari del doppi strato lipidico.
I carotenoidi possono avere accesso anche all’ambiente acquoso se in associazione a proteine, cui si legano in modo non covalente. Nel plasma sono trasportati dalle lipoproteine.
Colore
Il sistema di doppi legami coniugati, agendo come un cromoforo, è responsabile del colore dei carotenoidi. Tuttavia, perché il colore sia percettibile, devono essere presenti almeno sette doppi legami coniugati, per cui molecole come il fitofluene, che ha cinque legami coniugati, o il fitoene, che ha tre doppi legami coniugati, sono incolori.
All’aumentare del numero dei doppi legami coniugati il colore vira dal giallo, all’arancio, fino al rosso; ad esempio, la luteina, l’alfa-criptoxantina e la violaxantina sono gialle, l’alfa-carotene, il beta-carotene, e il gamma-carotene sono arancio, e il licopene è rosso.
Ruolo nelle piante
Nelle piante i carotenoidi svolgono molte funzioni.
Contribuiscono alla protezione della cellula nei confronti del danno ossidativo, aumentano l’assorbimento della luce solare, sono i precursori di fitormoni, come l’acido abscissico, e intervengono nelle vie di segnalazione.
Concorrono inoltre, con la clorofilla e gli antociani, una classe di flavonoidi, che sono il principale tipo di polifenoli, a determinare il colore delle foglie, dei frutti, delle verdure, dei cereali, colorazione che ha la funzione di attirare gli animali così da promuovere l’impollinazione e la dispersione dei semi.
Possono infine fungere anche da agenti repellenti per fitofagi e patogeni.
Quenching dell’ossigeno singoletto
Negli organismi che svolgono la fotosintesi ossigenica i carotenoidi concorrono alla protezione dei danni conseguenti all’eccessiva irradiazione solare.
Quando la luce che arriva complesso antenna del fotosistema II eccede la capacità di conversione dei centri di reazione, il surplus energetico può far si che la clorofilla permanga nel suo stato eccitato, il che può portare alla formazione del suo stato di tripletto. L’energia in eccesso può essere ceduta dalla clorofilla all’ossigeno molecolare determinando la formazione dell’ossigeno singoletto, una delle specie reattive dell’ossigeno, che è in grado di danneggiare l’apparato fotosintetico stesso e, più in generale, gli acidi nucleici, le proteine, gli acidi grassi insaturi di membrana, nonché i cofattori enzimatici.
I carotenoidi possono prevenire il danno ossidativo causato dall’ossigeno singoletto in due modi.
Possono reagire direttamente con l’ossigeno singoletto, un processo noto come quenching dell’ossigeno singoletto. In questo caso l’energia viene trasferita al carotenoide dall’ossigeno singoletto, che torna allo stato fondamentale di tripletto, mentre il carotenoide passa allo stato di tripletto, dissipando poi l’energia in eccesso in forma di calore. La capacità di disattivazione dell’ossigeno singoletto aumenta all’aumentare del numero dei doppi legami coniugati, divenendo massima da nove o più doppi legami coniugati.
Studi in vitro sembrano indicare che i carotenoidi monociclici siano in grado di disattivare l’ossigeno singoletto con maggiore efficienza rispetto alle strutture acicliche, e che la presenza di gruppi carbonilici, come nell’astaxantina, aumenti il potenziale antiossidante del carotenoide rispetto a quelli con gruppi idrossilici, probabilmente grazie al più esteso sistema di doppi legami coniugati. Sembra che ogni carotenoide, prima di incorrere in reazioni di degradazione, sia in grado di interagire e disattivare circa 1000 molecole di ossigeno singoletto.
I carotenoidi sono anche in grado di prevenire la formazione dell’ossigeno singoletto assorbendo l’energia della clorofilla eccitata, passando a loro volta in uno stato di tripletto. Essendo il loro livello energetico inferiore a quello dell’ossigeno singoletto, una volta formatisi decadono spontaneamente allo stato fondamentale.
Scavenging dei radicali liberi
I carotenoidi sono in grado di agire da scavenger dei radicali liberi, quali il radicale idrossilico, i radicali perossidici, il radicale superossido, o il radicale dell’ossido nitrico. La loro azione può avvenire secondo quattro modalità differenti.
Può verificarsi il trasferimento di un elettrone dal carotenoide al catione radicale, a formare il catione radicale del carotenoide, mentre il radicale diventa una specie neutra. Il catione del radicale carotenoide può essere rigenerato nel carotenoide di partenza grazie all’azione di altri antiossidanti cellulari come la vitamina E, la vitamina C e il glutatione.
Si può avere il trasferimento di un protone dal carotenoide al radicale, che diventa una specie neutra, mentre il carotenoide diviene un radicale anione.
La neutralizzazione del radicale può avvenire a seguito di reazioni di addizione, come nel caso della neutralizzazione dei radicali idrossilici e perossidici.
Un atomo di idrogeno può essere trasferito dal carotenoide al radicale, con formazione di un radicale carotenoide stabilizzato per risonanza.
Ruolo fotoassorbente
I carotenoidi sono presenti nei complessi antenna in qualità di pigmenti fotoassorbenti accessori, principalmente associati alle proteine antenna.
Grazie al sistema di doppi legami coniugati sono in grado di assorbire la luce solare nell’intervallo 400-500 nm. L’energia assorbita viene quindi convogliata ai centri di reazione.
Gli spettri di assorbimento dei singoli carotenoidi sono determinati dalla lunghezza della coniugazione e dal tipo di gruppi funzionali presenti.
Il principale carotene coinvolto nell’assorbimento della radiazione solare è il beta-carotene, presente nel nucleo del fotosistema I e II di tutti gli organismi che operano una fotosintesi ossigenica, mentre la luteina, la violaxantina, la neoxantina e la zeaxantina, le principali xantofille nelle piante, sono legate ai complessi di raccolta della luce.
Fonti alimentari
Gli esseri umani non sono in grado di sintetizzare i carotenoidi, li assumono dagli alimenti, li accumulano principalmente nel fegato e nel tessuto adiposo, ma anche nei polmoni, nei reni, nel cervello e nelle ossa, dove svolgono molte funzioni.
Di tutti i carotenoidi attualmente identificati, un centinaio sono presenti negli alimenti consumati dall’uomo, mentre, se si considera il singolo alimento, in genere ne sono presenti da uno a cinque principali, accompagnati da altri presenti in tracce o comunque piccole quantità.
Frutta e verdura sono la principale fonte di carotenoidi nella dieta umana. Tra le verdure, quelle a foglia verde sono ricche di beta-carotene, il carotenoide più abbondante nella dieta e nei tessuti umani, e luteina, seguiti da violaxantina e neoxantina, la radice delle carote è ricca di beta-carotene, la zucca di alfa-carotene, il pomodoro di licopene, e i peperoni di capsantina e capsorubina. I carotenoidi della frutta sono molto vari, e quelli dei frutti maturi possono essere diversi da quelli dei frutti acerbi. Inoltre, la maggior parte dei carotenoidi presenti nei frutti maturi sono esterificati con acidi grassi. Tuttavia, quelli di alcuni frutti, in particolare dei frutti che a maturazione terminata rimangono verdi, come il kiwi, subiscono una limitata o assente esterificazione.
Le uova di gallina, grazie al fatto che le galline ovaiole mangiano per la maggior parte il mais, possono essere una buona fonte di alcuni carotenoidi, come la luteina e zeaxantina, responsabili del colore giallo-arancio del tuorlo.
Infine, molluschi e crostacei, che con i pesci sono le fonti per eccellenza di acidi grassi polinsaturi omega-3 a catena lunga, quali l’acido eicosapentaenoico e l’acido docosaesaenoico, sono anche la principale fonte dei carotenoidi prodotti dalle microalghe e che raramente si trovano nelle piante, quali l’astaxantina presente nei gamberetti, ma anche nel salmone.
Additivi alimentari
I carotenoidi aggiunti ai cibi come additivi alimentari, a differenza dei carotenoidi naturalmente presenti, sono relativamente instabili, essendo suscettibili all’azione dell’ossigeno e della luce solare, all’auto-ossidazione, e alla dispersione nell’alimento, che ne facilita la degradazione. Inoltre, alla temperatura corporea e a quella di conservazione degli alimenti i carotenoidi aggiunti sono presenti in forma cristallina a causa del loro elevato punto di fusione; per ovviare a questo problema sono utilizzati in forma di emulsione olio in acqua.
Grazie al loro potere colorante sono utilizzati come coloranti in prodotti che, a causa della lavorazione o della conservazione tendono a perdere parte del loro colore naturale, o per uniformare il colore del prodotto, come nel caso delle bevande, dei succhi di frutta o degli insaccati, o per intensificarne il colore, migliorandone quindi l’aspetto.
Essendo precursori di composti responsabili del sapore e aroma di alcuni alimenti, possono essere utilizzati come esaltatori di sapidità.
Poiché alcuni carotenoidi sono precursori della vitamina A, sono utilizzati per fortificare gli alimenti.
Va infine sottolineato che, quando aggiunti agli alimenti, i carotenoidi non sono dei potenti antiossidanti.
Carotenoidi e salute dell’uomo
Nell’uomo i carotenoidi svolgono importanti funzioni.
Il ruolo più importante è quello di precursori della vitamina A, che da sola controlla l’espressione di quasi 700 geni. Non tutti i carotenoidi ne sono precursori; solo quelli che posseggono l’anello beta-ionone non sostituito possono essere convertiti nella vitamina, conversione che avviene a livello intestinale o nei tessuti di destinazione. Il più importante è il beta-carotene, che è la provitamina A per eccellenza e, avendo due anelli beta-ionone non sostituiti, viene convertito in due molecole di vitamina A. L’alfa-carotene e la beta-criptoxantina, avendo un solo anello beta-ionone non sostituito, possono dare una sola molecola di vitamina A, quindi hanno il 50% di attività di provitamina A.
L’importanza dell’attività di provitamina A svolta da alcuni carotenoidi è sottolineata dal fatto che la deficienza della vitamina può portare a cecità notturna e xeroftalmia, rallentare la crescita e la rigenerazione delle mucose, aumentare il tasso di mortalità causato dalle malattie infettive a seguito di disfunzioni della risposta immunitaria, ed è la singola causa più importante di cecità infantile nei paesi in via di sviluppo.
I carotenoidi contribuiscono alla salute dell’uomo anche concorrendo alla prevenzione di numerose patologie croniche attraverso meccanismi differenti rispetto all’azione di provitamina A, come l’azione antiossidante, antinfiammatoria e di fotoprotezione. Di seguito alcuni esempi.
E’ stato osservato che una supplementazione con luteina e zeaxantina, due xantofille che si accumulano nella macula lutea, è associata a un miglioramento della funzione visiva e a una riduzione del rischio di progressione della degenerazione maculare in tarda età.
Luteina e zeaxantina sembrano avere anche un effetto preventivo sullo sviluppo della cataratta e di retinopatie, e la luteina anche un effetto positivo sulle funzioni cognitive.
Tra i caroteni, il consumo di cibi ricchi di licopene è stato associato a una riduzione dell’incidenza di certi tipi di tumore, come il tumore della prostata e dello stomaco, e a una riduzione del rischio cardiovascolare.
Carotenoidi nel plasma
Alfa-carotene, beta-carotene, licopene, luteina, beta-criptoxantina e zeaxantina costituiscono oltre il 90% dei carotenoidi presenti nel plasma.
Le concentrazioni plasmatiche dei carotenoidi dipendono da diversi fattori tra cui i metodi di cottura dei cibi, la quantità di lipidi nella dieta, variabili individuali riguardanti i processi di digestione dei lipidi e assorbimento dei lipidi medesimi, con i sali biliari a svolgere un ruolo essenziale, e i livelli di vitamina A nell’organismo. Considerando quest’ultimo fattore, un soggetto che abbia bassi livelli di vitamina A avrà probabilmente tassi di conversione dei carotenoidi precursori della vitamina A più elevati, e ciò potrebbe riflettersi in una minore concentrazione plasmatica.
Bibliografia
Amengual J. Bioactive properties of carotenoids in human health. Nutrients 2019;11(10):2388. doi:10.3390/nu11102388
Archetti, M., Döring T.F., Hagen S.B., Hughes N.M., Leather S.R., Lee D.W., Lev-Yadun S., Manetas Y., Ougham H.J. Unravelling the evolution of autumn colours: an interdisciplinary approach. Trends Ecol Evol 2009;24(3):166-73. doi:10.1016/j.tree.2008.10.006
Böhm V., Lietz G., Olmedilla-Alonso B., Phelan D., Reboul E., Bánati D., Borel P., Corte-Real J., de Lera A.R., Desmarchelier C., Dulinska-Litewka J., Landrier J.F., Milisav I., Nolan J., Porrini M., Riso P., Roob J.M., Valanou E., Wawrzyniak A., Winklhofer-Roob B.M., Rühl R., Bohn T. From carotenoid intake to carotenoid blood and tissue concentrations – implications for dietary intake recommendations. Nutr Rev 2021;79(5):544-573. doi:10.1093/nutrit/nuaa008
Burrows T.L., Williams R., Rollo M., Wood L., Garg M.L., Jensen M., Clare Collins C.E. Plasma carotenoid levels as biomarkers of dietary carotenoid consumption: a systematic review of the validation studies. J Nutr Intermed Metab 2015;2(1-2):15-64. doi:10.1016/j.jnim.2015.05.001
de la Rosa L.A., Alvarez-Parrilla E., Gonzàlez-Aguilar G.A. Fruit and vegetable phytochemicals: chemistry, nutritional value, and stability. 1th Edition. Wiley J. & Sons, 2010
Desmarchelier C., Borel P. Overview of carotenoid bioavailability determinants: from dietary factors to host genetic variations. Trends Food Sci Technol 2017;69:270-280 doi:10.1016/j.tifs.2017.03.002
Eggersdorfer M., Wyss A. Carotenoids in human nutrition and health. Arch Biochem Biophys 2018;652:18-26. doi:10.1016/j.abb.2018.06.001
Gruszecki W.I., Strzałka K. Carotenoids as modulators of lipid membrane physical properties. Biochim Biophys Acta 2005;1740(2):108-15. doi:10.1016/j.bbadis.2004.11.015
Leyva-Porras C., Román-Aguirre M., Cruz-Alcantar, P., Pérez-Urizar, J.T., Saavedra-Leos M.Z. Application of antioxidants as an alternative improving of shelf life in foods. Polysaccharides 2021;2:594-607. doi:10.3390/polysaccharides2030036
Namitha K.K., Negi P.S. Chemistry and biotechnology of carotenoids. Crit Rev Food Sci 2010;50:728-760. doi:10.1080/10408398.2010.499811
Ramel F., Birtic S., Cuiné S., Triantaphylidès C., Ravanat J.L., Havaux M. Chemical quenching of singlet oxygen by carotenoids in plants. Plant Physiol 2012;158(3):1267-78. doi:10.1104/pp.111.182394
Rodriguez-Amaya D.B. Food carotenoids: chemistry, biology and technology. Wiley J. & Sons, 2015.
Saini R.K., Prasad P., Lokesh V., Shang X., Shin J., Keum Y.S., Lee J.H. Carotenoids: dietary sources, extraction, encapsulation, bioavailability, and health benefits – A review of recent advancements. Antioxidants 2022;11(4):795. doi:10.3390/antiox11040795
Sourkes T. L. The discovery and early history of carotene. Bull Hist Chem 2009;34(1)
Swapnil P., Meena M., Kumar Singh S., Praveen Dhuldhaj U., Harish, Marwal A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr Plant Biol 2021;26: 100203. doi:10.1016/j.cpb.2021.100203
Xu P., Chukhutsina V.U, Nawrocki W.J., Schansker G, Bielczynski L.W., Lu Y., Karcher D., Bock R., Croce R. Photosynthesis without β-carotene. eLife 2020;9:e58984. doi:10.7554/eLife.58984
Il carico di carboidrati è un’ottima strategia per ottimizzare le riserve energetiche nei muscoli prima dell’inizio di una competizione di resistenza come la maratona, l’ironman, il nuoto in acque libere o una gara di ciclismo su strada.
Cosa “mangia” il muscolo durante gli sport di resistenza?
Negli sport di resistenza le cause più probabili alla base dell’insorgenza della fatica sono la disidratazione e la deplezione dei carboidrati, in particolare del glicogeno muscolare ed epatico.
Per evitare la “crisi” dovuta alla deplezione dei carboidrati muscolari ed epatici è fondamentale avere alla partenza ottimi depositi di glicogeno.
Cosa influenza i depositi di glicogeno?
L’alimentazione nei giorni precedenti la gara.
Il livello di allenamento: chi è più allenato sintetizza più glicogeno e ha depositi potenzialmente maggiori perché ha enzimi più efficienti;
L’attività il giorno della gara e i giorni precedenti, in quanto se il muscolo non lavora non perde glicogeno. Quindi nei giorni che precedono la gara è bene fare allenamenti leggeri, così da non intaccarne le riserve, e curare l’alimentazione.
L’origine “svedese” del carico di carboidrati
Negli eventi che durano più di 90 minuti, avere riserve muscolari di glicogeno molto elevate (si parla di supercompensazione del glicogeno) può migliorare la performance, ossia il tempo necessario per completare una data distanza, di un 2-3% in confronto con una situazione in cui le riserve di glicogeno sono normali o basse. Nelle competizioni con durata inferiore ai 90 minuti i benefici della supercompensazione sembrano essere piccoli o assenti.
Gli atleti ben allenati possono ottenere la supercompensazione delle riserve di glicogeno anche senza ricorrere alla fase di deplezione dei carboidrati precedente al carico degli stessi, vecchia tecnica messa a punto da due ricercatori svedesi, Saltin e Hermansen, negli anni ’60 del secolo scorso.
I due ricercatori scoprirono che la concentrazione muscolare del glicogeno poteva essere raddoppiata seguendo nei sei giorni precedenti la gara una dieta di questo tipo:
tre giorni di dieta ipoglucidica (poverissima di carboidrati);
tre giorni di dieta iperglucidica, il cosiddetto carico di carboidrati (dieta ricchissima di carboidrati).
Questa dieta crea un sacco di problemi: i primi tre giorni senza carboidrati, ossia senza pasta, riso, pane, patate, legumi, frutta ecc., sono durissimi, ci possono essere anche sintomi simili alla depressione dovuti al carente apporto di glucosio al cervello, mentre i vantaggi che si ottengono sono pochi.
Inoltre, con le tecniche di preparazione attuali, tipo e quantità di lavoro svolto, già si riescono a ottenere livelli di glicogeno elevati, oltre i 2,5 g/kg.
Il carico di carboidrati “moderno”
Immaginando di avere la gara la domenica un possibile schema per ottenere la supercompensazione delle riserve di glicogeno può essere il seguente:
mercoledì, ossia 4 giorni prima della competizione, allenamento discreto e poi cena senza carboidrati;
da giovedì, quindi 3 giorni prima della competizione, dieta iperglucidica, ossia il carico di carboidrati e allenamenti leggeri.
Carico di Carboidrati: Dieta da 2500 kcal
La quantità di carboidrati necessaria per ripristinare le scorte di glicogeno o per promuoverne il carico varia in funzione della durata e dell’intensità del programma di allenamento, ed è compresa tra 5 e 12 g/kg/d a seconda dell’atleta e della sua attività. Con apporti di carboidrati maggiori si possono ottenere scorte più elevate di glicogeno, ma non sempre questo determina prestazioni migliori. Inoltre c’è anche da considerare il fatto che l’accumulo di glicogeno si accompagna a un aumento di peso dovuto alla ritenzione di acqua, circa 3 grammi di acqua per ogni grammo di glicogeno, e per alcuni sport questo potrebbe non essere vantaggioso.
Bibliografia
Burke L.M., Hawley J.A., Wong S.H.S., & Jeukendrup A. Carbohydrates for training and competition. J Sport Sci 2011;29:Sup1,S17-S27. doi:10.1080/02640414.2011.585473
Hargreaves M., Hawley J.A., & Jeukendrup A.E. Pre-exercise carbohydrate and fat ingestion: effects on metabolism and performance. J Sport Sci 2004;22:31-38. doi10.1080/0264041031000140536
Jeukendrup A.E., C. Killer S.C. The myths surrounding pre-exercise carbohydrate feeding. Ann Nutr Metab 2010;57(suppl 2):18-25. doi:10.1159/000322698
Moseley L., Lancaster G.I, Jeukendrup A.E. Effects of timing of pre-exercise ingestion of carbohydrate on subsequent metabolism and cycling performance. Eur J Appl Physiol 2003;88:453-458. doi:10.1007/s00421-002-0728-8
L’ipotesi alla base della dieta alcalina afferma gli alimenti proteici e i cereali, con un basso apporto di potassio, portano ad una dieta acida, a una escrezione acida netta a livello renale, a un aumento del calcio nelle urine, e al rilascio di calcio dallo scheletro: tutto ciò causerebbe osteoporosi.
E’ vero?
Il calcio presente nelle ossa in forma di carbonati e fosfati rappresenta un grande serbatoio di basi, cioè di sostanze con cui tamponare l’acidità nel corpo. In risposta a un carico acido, come quello derivante da diete ricche di proteine, questi sali vengono rilasciati in circolo per mantenere costante il pH. Il minerale è quindi eliminato nelle urine. E’ stato stimato che la quantità persa da un soggetto che segua questo tipo di diete potrebbe arrivare a circa 480 g in 20 anni o quasi metà della massa scheletrica di calcio!
Anche se queste perdite di calcio possono essere tamponate mangiando alimenti ricchi di sostanze alcaline, come frutta e verdura, e molte informazioni on-line così come un certo numero di libri promuovono una dieta alcalina per la salute delle ossa, una recente meta-analisi ha mostrato che l’associazione causale tra malattia dell’osso da osteoporosi e carico acido dietetico non è supportata da prove e non vi è alcuna evidenza che una dieta alcalina sia protettiva nei confronti della salute delle ossa, mentre protegge contro il rischio di calcoli renali.
Nota: è possibile che la frutta e la verdura siano benefiche per la salute delle ossa attraverso meccanismi diversi da quanto supposto dall’ipotesi della dieta alcalina.
Qual è il ruolo delle proteine?
L’eccesso di proteine nella dieta, a causa del loro elevato carico acido renale, può ridurre la densità ossea se non tamponato dalla ingestione di alimenti che ricchi di alcali, cioè frutta e verdura. Tuttavia, un adeguato apporto di proteine è necessario per il mantenimento dell’integrità ossea. Pertanto, potrebbe essere necessario l’aumento della quantità di frutta e verdura, invece della eccessiva riduzione delle proteine.
E’ quindi consigliabile consumare una dieta normo-proteica ricca di frutta e verdura e povera di sodio, come la dieta mediterranea. Un corretto abbinamento dei cibi prevede l’ingestione di alimenti con un carico acido negativo accompagnati da altri un carico acido positivo. Esempio: pasta più verdure o carne più verdura e frutta, le ultime due, in particolare la verdura, in porzioni generose.
Dieta alcalina e massa muscolare
Con l’avanzare dell’età vi è una perdita di massa muscolare che predispone a cadute e fratture. Una dieta ricca di potassio, ottenuto da frutta e verdura, così come un carico acido ridotto, protegge la massa muscolare sia negli uomini che nelle donne anziane.
Dieta alcalina e ormone della crescita
Nei bambini gravi forme di acidosi metabolica sono associate a bassi livelli di ormone della crescita, con conseguente bassa statura: la correzione dell’acidosi con bicarbonato o citrato di potassio aumenta in modo significativo l’ormone della crescita migliorando la crescita.
Nelle donne in postmenopausa, l’uso di sufficiente bicarbonato di potassio nella dieta per neutralizzare il carico acido netto giornaliero ha determinato un aumento significativo dell’ormone della crescita e dell’osteocalcina risultante.
Migliorare i livelli di ormone della crescita può ridurre i fattori di rischio cardiovascolare, migliorare la qualità della vita, la composizione corporea, e anche la memoria e la cognizione.
Conclusioni
La dieta alcalina può comportare un certo numero di benefici per la salute.
L’aumento del consumo di frutta e verdura migliorerebbe il rapporto potassio/sodio e ciò potrebbe essere di beneficio per la salute delle ossa, potrebbe ridurre la perdita muscolare, così come mitigare altre malattie croniche come l’ipertensione e ictus.
L’aumento dell’ormone della crescita può avere molti effetti positivi, dalla salute cardiovascolare alla memoria e cognizione.
L’aumento del magnesio intracellulare è un altro vantaggio ulteriore della dieta alcalina; ad esempio, il magnesio, necessario per attivare la vitamina D, comporterebbe numerosi benefici aggiunti nei sistemi ormonali in cui la vitamina è implicata.
Va notato che una delle prime considerazioni riguardanti una dieta alcalina, che comprende più frutta e verdura, è sapere in che tipo di terreno questi cibi sono state coltivati poiché può influenzare in modo significativo il loro contenuto in minerali e quindi il loro potere tamponante.
Bibliografia
Fenton T.R., Lyon A.W., Eliasziw M., Tough S.C., Hanley D.A. Meta-analysis of the effect of the acid-ash hypothesis of osteoporosis on calcium balance. J Bone Miner Res 2009;24(11):1835-1840. doi:10.1359/jbmr.090515
Fenton T.R., Lyon A.W., Eliasziw M., Tough S.C., Hanley D.A. Phosphate decreases urine calcium and increases calcium balance: a meta-analysis of the osteoporosis acid-ash diet hypothesis. Nutr J 2009;8:41. doi:10.1186/1475-2891-8-41
Fenton T.R., Tough S.C., Lyon A.W., Eliasziw M., Hanley D.A. “Causal assessment of dietary acid load and bone disease: a systematic review and meta-analysis applying Hill’s epidemiologic criteria for causality.” Nutr J 2011;10:41. doi:10.1186/1475-2891-10-41
Schwalfenberg G.K. The alkaline diet: is there evidence that an alkaline pH diet benefits health? J Environ Public Health 2012; Article ID 727630. doi:10.1155/2012/727630
La vita dipende da adeguati valori di pH sia all’interno dell’organismo e delle cellule che lo compongono che nell’ambiente circostante.
Gli esseri umani per evitare l’acidosi metabolica e sopravvivere necessitano di uno strettissimo controllo del valore del pH del sangue, che è pari a circa 7,4. A titolo di confronto, negli ultimi 100 anni il pH del mare è sceso da 8,2 a 8,1 a causa della crescente dissoluzione al suo interno di anidride carbonica, con un impatto negativo sulla vita degli oceani.
Anche il contenuto in minerali del cibo che mangiamo è notevolmente influenzato dal pH del terreno in cui le piante sono coltivate.
La scala del pH
Il pH ideale del terreno per ottenere la migliore disponibilità complessiva di nutrienti essenziali è compresa tra 6 e 7: un terreno acido, con pH inferiore a 6, può avere un ridotto contenuto in magnesio e calcio, mentre valori superiori a pH 7 possono rendere chimicamente non disponibili zinco, ferro, rame e manganese.
Acidosi metabolica e rivoluzione agricola e industriale
Nella dieta umana, dalla società dei cacciatori-raccoglitori alla presente, c’è stato un notevole cambiamento nel pH e nel carico acido netto della dieta umana. Con la rivoluzione agricola, e ancora più di recente, con l’industrializzazione, si è osservato:
un incremento della quantità di sodio rispetto a quella di potassio, con il rapporto potassio/sodio che si è invertito passando da 10 a 1 a 1 a 3, e del cloruro rispetto al bicarbonato;
Ne risulta una dieta che può portare ad acidosi metabolica, una condizione che non corrisponde a quelle che sono le esigenze nutrizionali geneticamente determinate.
Inoltre, con l’invecchiamento, si assiste a una progressiva perdita della funzione regolatrice renale dell’equilibrio acido-base e al conseguente aumento dell’acidosi metabolica indotto dalla dieta.
Infine, una dieta povera di carboidrati ma ad alto contenuto proteico, con il suo aumentato carico acido, determina cambiamenti molto piccoli nella chimica del sangue, e nei valori del suo pH, ma causa molti cambiamenti nella chimica delle urine. Si osserva infatti un aumento del calcio, dell’acido urico indissociato e del fosfato, mentre si riducono il magnesio, il citrato e il pH delle urine.
Tutto ciò aumenta il rischio di calcoli renali.
Il pH come scudo acido
Il corpo umano ha una straordinaria capacità di mantenere un pH stabile nel sangue, con i principali meccanismi di compensazione presenti a livello renale e respiratorio.
Il pH nel corpo varia notevolmente da una zona all’altra.
L’acidità massima si trova nello stomaco (pH 1,35-3,5), dove aiuta nella digestione e ci protegge nei confronti dei microrganismi opportunisti.
La pelle è abbastanza acida (pH 4-6,5), e questo fornisce un “scudo” acido che funge da barriera protettiva verso l’ambiente nei confronti della proliferazione microbica. Ciò si osserva anche nella vagina, dove un pH inferiore a 4,7 protegge dalla proliferazione microbica.
L’urina ha un pH variabile da acido ad alcalino a seconda delle necessità di equilibrare l’ambiente interno.
Neutralizza l’acidità gastrica, aiuto nella digestione
Succo pancreatico
8,8
Neutralizza l’acidità gastrica, aiuto nella digestione
Fluido vaginale
<4,7
Limita la sovracrescita dei microbi opportunistici
Liquido cerebrospinale
7,3
Bagna l’esterno del cervello
Liquido intracellulare
Tra 6,0 e 7,2
Dovuto alla produzione di acido nelle cellule
Siero venoso
7,35
Strettamente regolato
Siero arterioso
7,4
Strettamente regolato
Modificato da: Schwalfenberg G.K.; vedere nella Bibliografia
Bibliografia
Fenton T.R., Lyon A.W., Eliasziw M., Tough S.C., Hanley D.A. Meta-analysis of the effect of the acid-ash hypothesis of osteoporosis on calcium balance. J Bone Miner Res 2009;24(11):1835-1840. doi:10.1359/jbmr.090515
Fenton T.R., Lyon A.W., Eliasziw M., Tough S.C., Hanley D.A. Phosphate decreases urine calcium and increases calcium balance: a meta-analysis of the osteoporosis acid-ash diet hypothesis. Nutr J 2009;8:41. doi:10.1186/1475-2891-8-41
Fenton T.R., Tough S.C., Lyon A.W., Eliasziw M., Hanley D.A. Causal assessment of dietary acid load and bone disease: a systematic review and meta-analysis applying Hill’s epidemiologic criteria for causality. Nutr J 2011;10:41. doi:10.1186/1475-2891-10-41
Schwalfenberg G.K. The alkaline diet: is there evidence that an alkaline pH diet benefits health? J Environ Public Health 2012; Article ID 727630. doi:10.1155/2012/727630
La letteratura scientifica internazionale è concorde nello stabilire come limite inferiore per l’apporto calorico giornaliero 1200 kcal per la donna e 1500 kcal per l’uomo.
Per rendere il bilancio calorico giornaliero negativo, perdere peso, ma soprattutto perdere massa grassa, alla valutazione delle reali necessità caloriche del soggetto sarà da affiancarsi:
la corretta suddivisione dei pasti nella giornata;
un’aumentata attività fisica, grazie alla quale il bilancio negativo potrà essere ottenuto senza particolari sacrifici a tavola.
Bilancio Calorico Giornaliero
Questo renderà il dimagrimento più facile e proteggerà dai successivi aumenti di peso, in particolare da quello che è conosciuto come effetto yo-yo.
In definitiva ci deve essere un cambio nello stile di vita.
Dunque, la strategia migliore per perdere massa grassa non è una drastica riduzione dell’apporto calorico e neppure seguire regimi alimentari costrittivi, ad es. diete che promettono miracoli come la “dieta del minestrone”, la dieta Plank, la dieta Master Cleanse e molte altre che obbligano a eliminare o ridurre fortemente l’apporto di determinati macronutrienti, quasi sempre i carboidrati, con apporti spesso esagerati di proteine. Condotte simili possono essere:
molto stressanti dal punto di vista psicologico;
non percorribili per lunghi periodi;
rischiose per la salute a causa di inevitabili carenze di nutrienti.
Infine non assicurano affatto che i chili persi siano solo o quasi solo massa grassa, e spesso sono seguite da aumenti del peso corporeo sostanziali ossia da quello che è conosciuto come effetto yo-yo.
Perché?
Effetti sulla massa grassa della restrizione calorica
Una riduzione eccessiva dell’apporto calorico vuol dire mangiare molto poco. Ciò determina il rischio, elevato, di non assumere in quantità adeguate i diversi nutrienti essenziali, quelli cioè che non possiamo sintetizzare, come le vitamine, alcuni aminoacidi, gli acidi grassi essenziali, che sono una sottoclasse di lipidi, e i minerali, tra cui il calcio, indispensabile per il metabolismo osseo in ogni fase della vita, o il ferro che è utilizzato in molte funzioni del nostro organismo come il trasporto di ossigeno ai tessuti. Tutto ciò si traduce anche in una depressione del metabolismo e quindi una riduzione dei consumi. Se poi la riduzione dell’apporto calorico è eccessiva o addirittura ci sono periodi di digiuno al danno si aggiunge la beffa in quanto si perderà una quota di massa magra. In che modo?
Riduzione delle calorie e ruolo dei carboidrati
Il glucosio rappresenta l’unica fonte di energia per i globuli rossi e alcune zone del cervello, mentre altre zone possono ricavare energia anche dai corpi chetonici, un prodotto del metabolismo degli acidi grassi.
A riposo il cervello estrae il 10% del glucosio dal circolo, una quantità non trascurabile, circa 75 mg/min., se si considera che il suo peso è di circa 1,5 kg. Per mantenere costante la glicemia, e quindi assicurare un costante rifornimento di glucosio ai vari tessuti, è necessario assumere carboidrati, o in alternativa aminoacidi, entrambe facilmente ottenibili dagli alimenti.
In caso di un apporto dietetico di carboidrati scarso o assente, considerando che dopo circa 18 ore si esaurisce il glicogeno epatico che rilascia in circolo glucosio, l’organismo sintetizza de novo glucosio a partire da alcuni aminoacidi attraverso un processo detto gluconeogenesi. Questa via metabolica è attiva anche dopo un pasto normale, ma in caso di digiuno la sua importanza aumenta.
Ma qual è la principale fonte di aminoacidi nell’organismo nel caso in cui anche il loro apporto dietetico sia scarso o assente? Le proteine endogene, ed esiste una sorta di gerarchia nella loro utilizzazione cioè prima si consumano quelle che servono di meno e solo in seguito quelle più importanti. Per primi saranno utilizzati gli enzimi della digestione, pepsina, chimotripsina, elastasi, carbossipeptidasi e aminopeptidasi, in tutto 35-40 g; in seguito fegato e pancreas rallentano la loro attività di sintesi di proteine da esportare e gli aminoacidi inutilizzati sono avviati verso la gluconeogenesi. E’ evidente che queste riserve di aminoacidi sono abbastanza modeste, e sarà il muscolo a farsi carico di fornirne le quantità necessarie, cioè ha inizio la proteolisi delle proteine del muscolo.
Da notare che comunque non esiste una sequenzialità assoluta nella degradazione delle diverse proteine, c’è invece un intreccio in cui, procedendo, certe vie perdono di importanza ed altre ne acquistano.
Quindi per mantenere la glicemia costante viene ridotta la componente proteica del muscolo, compreso quello scheletrico, un tessuto che rappresenta una discreta quota del valore del metabolismo basale e che con l’attività fisica è in grado di aumentare considerevolmente il suo dispendio energetico: dunque fondamentale ai fini della perdita di peso, cioè di massa grassa, e del successivo mantenimento. E’ come se si riducesse la cilindrata del motore.
Una cosa a cui non si pensa è che anche il cuore è un muscolo per cui potrà essere soggetto agli stessi processi visti per il muscolo scheletrico.
In definitiva produrre glucosio a partire dalle proteine, anche di origine alimentare, è come scaldarsi al camino bruciando il mobilio del settecento, gli aminoacidi, avendo a disposizione legna da ardere, i carboidrati alimentari.
Pertanto un adeguato apporto di carboidrati con l’alimentazione previene la perdita eccessiva delle proteine ossia c’è un effetto di risparmio delle proteine svolto dai carboidrati.
Nota: i mammiferi, e quindi gli esseri umani, non hanno la capacità di sintetizzare glucosio a partire dai grassi.
Carboidrati: cosa entra quando loro escono
L’eliminazione o la forte riduzione dell’apporto di carboidrati con la dieta si traduce in un aumentato apporto di proteine, lipidi, incluso il colesterolo, in quanto sarà aumentata l’assunzione di prodotti di origine animale, uno dei principali difetti delle diete iperproteiche. Infatti, nell’organismo non esistono riserve di aminoacidi per cui questi vengono metabolizzati e, come sottoprodotto della loro utilizzazione, si forma ammoniaca che dovrà essere eliminata in quanto tossica.
Per questo motivo le diete iperproteiche comportano un lavoro extra per fegato e reni, e anche per questo non sono esenti da potenziali rischi per la salute.
Un aumentato apporto di grassi molto spesso si traduce in un aumento dell’assunzione di acidi grassi saturi, acidi grassi trans e colesterolo, con tutte le conseguenze che ciò può avere a livello cardiovascolare.
Quanto detto non deve incitare ad assumere quantità elevate di carboidrati. Questa classe di macronutrienti dovrebbe rappresentare il 55-60% della quota calorica giornaliera, i grassi il 25-30% (olio di olivain primis) e la restante quota alle proteine: dunque una composizione in macronutrienti che si rifà alla dieta prudente o alla dieta mediterranea.
Massa grassa e ingresso nella fase di carestia/malattia
Una riduzione eccessiva dell’apporto calorico viene registrata a livello dei nostri meccanismi di difesa come un “ingresso” in una fase di carestia/malattia.
L’abbondanza di cibo è una caratteristica della nostra epoca, almeno nei paesi industrializzati, mentre il nostro organismo si è evoluto nel corso di centinaia di migliaia di anni durante i quali non c’era l’attuale abbondanza: dunque è stato programmato per cercare di superare con il minimo dei danni periodi di carestia. Se l’apporto calorico viene ridotto drasticamente si mima una carestia: quello che l’organismo fa è di abbassare i consumi, ridurre il metabolismo basale ossia consuma di meno e quindi anche mangiando poco non si otterranno grandi perdite di massa grassa. E’ come se a una macchina si abbassasse la cilindrata, consumerà meno, e nel nostro caso brucerà meno massa grassa.
Effetto yo-yo
Il weight cycling o effetto yo-yo, ossia fasi ripetute di perdita e acquisto di peso, appare correlato a un eccesso di peso e a un accumulo di grasso a livello addominale.
Diversi studi suggeriscono l’esistenza nelle donne di un legame tra effetto yo-yo e diverse condizione quali:
un aumento importante del binge eating disorder o sindrome da alimentazione incontrollata;
un senso di depressione riguardo al peso
Infine l’effetto yo-yo si associa anche a una maggiore facilità di prendere peso rispetto alle donne che non sono soggette. A questo riguardo c’è da sottolineare che questa condizione si verifica nell’arco di anni durante i quali, invecchiando, la velocità del metabolismo inevitabilmente tende a diminuire: questo potrebbe rendere più difficoltose le perdite successive.
Bibliografia
Cereda E., Malavazos A.E., Caccialanza R., Rondanelli M., Fatati G. and Barichella M. Weight cycling is associated with body weight excess and abdominal fat accumulation: a cross-sectional study. Clin Nutr 2011;30(6):718-723. doi:10.1016/j.clnu.2011.06.009
Montani J-P., Viecelli A.K., Prévot A. & Dulloo A.G. Weight cycling during growth and beyond as a risk factor for later cardiovascular diseases: the ‘repeated overshoot’ theory. Int J Obes (Lond) 2006;30:S58-S66. doi:10.1038/sj.ijo.0803520
Ravussin E., Lillioja S., Knowler W.C., Christin L., Freymond D., Abbott W.G.H., Boyce V., Howard B.V., and Bogardus C. Reduced rate of energy expenditure as a risk factor for body-weight gain. N Engl J Med 1988;318:467-472. doi:10.1056/NEJM198802253180802
Sachiko T. St. Jeor S.T. St., Howard B.V., Prewitt T.E., Bovee V., Bazzarre T., Eckel T.H., for the AHA Nutrition Committee. Dietary Protein and Weight Reduction. A Statement for Healthcare Professionals From the Nutrition Committee of the Council on Nutrition, Physical Activity, and Metabolism of the American Heart Association. Circulation 2001;104:1869-1874. doi:10.1161/hc4001.096152
Il glicogeno muscolare è un’importante riserva di energia negli esercizi prolungati di intensità medio-alta, importanza che aumenta nel caso di esercizi intervallati di alta intensità, comuni negli allenamenti di nuotatori, corridori, vogatori e negli sport di squadra, o in lavori di resistenza contro pesi. Se ad esempio si considera la maratona, circa l’80% dell’energia necessaria deriva dall’ossidazione dei carboidrati, per la maggior parte glicogeno muscolare.
La deplezione dei livelli di glicogeno muscolare è correlata con l’insorgenza della fatica, sebbene non siano ancora chiari i meccanismi molecolari alla base di questa relazione. Una delle ipotesi è che esista una concentrazione minima di glicogeno che viene “protetta” ed è resistente all’utilizzo durante l’esercizio, forse per assicurare una riserva di energia in caso di estrema necessità.
Data la stretta relazione tra deplezione del glicogeno muscolare e fatica, la sua velocità di ripristino nel post-esercizio è uno dei fattori più importanti nel determinare il tempo necessario al recupero.
Infine l’atleta maggiormente allenato non solo ha depositi di glicogeno muscolare potenzialmente maggiori, ma è anche in grado di sintetizzarlo più velocemente grazie a enzimi più efficienti.
Per produrre glicogeno è indispensabile assumere carboidrati; ma quanti, quali, quando e con che frequenza?
Nel tentativo di ripristinare il più velocemente possibile le riserve di glicogeno muscolare è utile conoscere l’andamento bifasico che può assumere la sua velocità di sintesi a seguito di allenamenti o gare che comportino deplezioni delle sue riserve muscolari pari ad almeno il 75% del valore a riposo e non a digiuno.
Conoscere e quindi sfruttare la tale bifasicità è importante per quegli atleti che siano impegnati in più allenamenti giornalieri, o che abbiano poco tempo a disposizione per il recupero tra un allenamento impegnativo e il successivo, ossia meno di 8 ore, al fine di massimizzarne la sintesi e ottenere la migliore performance nella sessione successiva.
Le due fasi sono caratterizzate da:
una diversa sensibilità all’insulina circolante;
una diversa velocità di sintesi.
La prima fase
La prima fase, immediatamente successiva al termine dell’attività e della durata di 30-60 minuti, è insulino-indipendente, ossia l’uptake del glucosio da parte della cellula muscolare come la sintesi della molecola al suo interno sono indipendenti dall’azione dell’ormone.
Questa fase è caratterizzata da una elevata velocità di sintesi che però si riduce rapidamente se non si assumono carboidrati: la massima velocità si registra nei primi 30 minuti, per poi ridursi a circa 1/5 dal 60° minuto, e a circa 1/9 al 120° minuto dal termine dell’esercizio.
Come è possibile sfruttare questa prima fase per ripristinare quanto più possibile le scorte muscolari di glicogeno? Facendo si che al muscolo arrivi la maggior quantità possibile di glucosio nei momenti immediatamente successivi al termine dell’attività, meglio se entro i primi 30 minuti.
Cosa assumere?
Carboidrati a alto indice glicemico, ma di facile digestione e assorbimento.
E’ quindi consigliabile sostituire cibi, magari anche ad alto indice glicemico, che necessitano di un certo tempo per la digestione e il successivo assorbimento, con soluzioni/gel contenenti ad esempio glucosio e/o saccarosio. Queste soluzioni/gel assicurano la massima velocità possibile di assorbimento e rifornimento di glucosio al muscolo in quanto contengono solo glucosio e sono prive di fibre o altro che rallenterebbero la digestione dei carboidrati e il successivo assorbimento dei monosaccaridi, sono cioè in grado di produrre elevate glicemie in un tempo relativamente breve.
Da sottolineare ulteriormente che il ricorso nell’immediato post-esercizio a tali soluzioni/gel contenenti carboidrati a rapida disponibilità è consigliabile solo quando il tempo di recupero tra un esercizio che causa una forte deplezione del glicogeno muscolare e il successivo è breve, meno di 8 ore.
Sarà possibile giocare anche sulla temperatura e concentrazione della soluzione per accelerarne il transito gastrico.
Assumere carboidrati, ma in che quantità?
Sono stati condotti molti studi per cercare di definire la quantità ideale di carboidrati da assumere.
Se nel post-esercizio l’atleta non si alimenta la velocità di sintesi del glicogeno è molto bassa, mentre se immediatamente dopo il termine del lavoro assume quantità adeguate di carboidrati la velocità può raggiungere valori oltre 20 volte maggiori.
Dal confronto della letteratura sembra ragionevole affermare che, a seguito di allenamenti che riducano le scorte di glicogeno muscolare a valori inferiori al 75% dei valori a riposo e non a digiuno, la massima velocità di sintesi si ottenga con assunzioni di carboidrati, ad alto indice glicemico ed elevata velocità di digestione e assorbimento, pari a circa 1,2 g/kg di peso corporeo/h per le 4-5 ore successive dal termine dell’esercizio stesso.
In questo modo si determina la produzione di una quantità di glicogeno maggiore del 150% rispetto all’ingestione di 0,8 g/kg/h.
Poiché aumenti fino a 1,6 g/kg/h non hanno portato ulteriori incrementi, la quantità di carboidrati pari a 1,2 g/kg/h può essere considerata quella ottimale per massimizzare la velocità di risintesi delle scorte di glicogeno muscolare nel post-esercizio.
Con che frequenza?
Riguardo alla frequenza di assunzione è stato osservato che se i carboidrati sono assunti di frequente, ogni 15-30 minuti, sembra ci sia un’ulteriore stimolazione dell’uptake del glucosio da parte del muscolo, come della ricostituzione del glicogeno muscolare rispetto ad assunzioni a intervalli di due ore. In particolare, le assunzioni nelle prime ore del post-esercizio sembrano ottimizzare livelli di glicogeno.
La seconda fase
La seconda fase delle sintesi del glicogeno muscolare ha inizio dalla fine della prima, perdura sino all’inizio del pasto precedente l’impegno successivo, dunque da alcune ore a giorni, ed è insulino-dipendente, ossia l’uptake del glucosio da parte della cellula muscolare come la sintesi del glicogeno al suo interno sono sensibili ai livelli circolanti dell’ormone.
Inoltre si osserva una significativa riduzione della velocità di sintesi del glicogeno muscolare: con un’assunzione adeguata di carboidrati la velocità si attesta su valori inferiori di circa il 10-30% rispetto a quelli della prima fase.
Questa fase può perdurare per diverse ore, ma tende a essere più breve se:
l’apporto di carboidrati è elevato;
la sintesi del glicogeno è più attiva;
i livelli di glicogeno muscolare sono aumentati.
Come è possibile sfruttare questa fase per ottimizzare la velocità di sintesi del glicogeno muscolare?
Le evidenze sperimentali indicano che pasti con carboidrati ad alto indice glicemico sono più efficaci di quelli con carboidrati a basso indice glicemico. Ma se tra un allenamento e il successivo passano giorni e non ore, non ci sono evidenze a favore di carboidrati ad alto indice glicemico rispetto a quelli a basso indice purché ne sia assunta una quantità adeguata.
Velocità di sintesi del glicogeno e assunzione di carboidrati e proteine
La contemporanea assunzione di carboidrati e proteine, o aminoacidi insulino-tropici liberi, permette di ottenere velocità di sintesi del glicogeno nel post-esercizio che non differiscono significativamente da quelle raggiunte con quantità maggiori di soli carboidrati. Questo potrebbe essere un vantaggio per l’atleta che ne potrà assumere quantità più contenute, limitando così l’insorgenza di eventuali complicazioni gastrointestinali comuni durante l’allenamento/gara dopo un loro consumo elevato.
Dall’analisi della letteratura sembra ragionevole affermare che, dopo un esercizio che comporti la deplezione di almeno il 75% delle riserve muscolari di glicogeno, si possano ottenere velocità di sintesi del glicogeno analoghe a quelle raggiunte con 1,2 g/kg/h di soli carboidrati, le maggiori ottenibili, con la coingestione di 0,8 g/kg/h di carboidrati e 0,4 g/kg/h di proteine, mantenendo le stesse tempistiche di ingestione, ossia ogni 15-30 minuti per le prime 4-5 ore del post-esercizio.
Le due fasi: meccanismi molecolari
In entrambe le fasi l’aumento della sintesi del glicogeno è conseguenza di un aumento:
della velocità di trasporto del glucosio nella cellula;
dell’attività della glicogeno sintetasi, l’enzima che catalizza la sintesi del glicogeno.
Tuttavia i meccanismi molecolari che sottostanno a queste modificazioni sono differenti.
Nella prima fase l’aumento della velocità di trasporto del glucosio, indipendente dalla presenza dell’insulina, è mediato dalla traslocazione, indotta dalla contrazione, dei trasportatori del glucosio, detti GLUT4, sulla membrana plasmatica della cellula muscolare.
In aggiunta, anche i bassi livelli di glicogeno agiscono da stimolo al trasporto in quanto si ritiene che gran parte delle vescicole contenenti il trasportatore siano legate al glicogeno, e dunque potrebbero divenire disponibili quando i suoi livelli sono ridotti.
Infine bassi livelli di glicogeno muscolare vanno anche a stimolare l’attività della glicogeno sintetasi: è stato dimostrato che il livello del glicogeno muscolare è un regolatore dell’attività dell’enzima molto più potente di quanto sia l’insulina.
Nella seconda fase l’aumento della sintesi è dovuto all’azione dell’insulina sui trasportatori del glucosio e sull’attività della glicogeno sintetasi della cellula muscolare. Questa sensibilità all’azione dell’insulina circolante, che può persistere per 48 ore a seconda dell’assunzione di carboidrati e della quantità di glicogeno muscolare risintetizzato, ha suscitato grande attenzione: è infatti possibile, tramite opportuni interventi nutrizionali, incrementarne la secrezione al fine di migliorare la sintesi del glicogeno stesso ma anche l’anabolismo proteico, riducendo al contempo la velocità di degradazione delle proteine stesse.
Insulina e velocità di sintesi del glicogeno muscolare
La contemporanea assunzione di carboidrati e proteine, o aminoacidi liberi, aumenta la secrezione di insulina postprandiale rispetto ai soli carboidrati; in alcuni studi sono stati osservati incrementi nella secrezione dell’ormone di 2-3 volte rispetto ai soli carboidrati.
E’ stato supposto che, data la maggior quantità di insulina circolante, si potessero ottenere ulteriori aumenti della velocità di sintesi del glicogeno rispetto a quelli osservati con i soli carboidrati, ma in realtà non sembra essere così. Se infatti la quantità di carboidrati viene portata a 1,2 g/kg/h, più 0,4 g/kg/h di proteine, non si osservano ulteriori aumenti nella velocità di sintesi se paragonati a quelli ottenuti con l’ingestione dei soli carboidrati nella stessa quantità, ossia 1,2 g/kg/h, che come detto, al pari della coingestione di 0,8 g/kg/h di carboidrati e 0,4 g/kg/h di proteine, danno la massima velocità raggiungibile nel post-esercizio, o in quantità isoenergetica, quindi 1,6 g/kg.
Insulina e accumulo preferenziale dei carboidrati
I livelli più elevati di insulina circolante raggiunti con la coingestione di carboidrati e proteine, o aminoacidi liberi, potrebbero stimolare un accumulo dei carboidrati ingeriti nei tessuti maggiormente sensibili all’azione dell’ormone, quali il fegato e il muscolo che ha precedentemente lavorato.
In questo modo si verificherebbe un loro deposito più efficiente ai fini dell’attività sportiva, in quanto i carboidrati verrebbero accumulati preferenzialmente anche nel muscolo, dove saranno in seguito utilizzati.
Bibliografia
Beelen M., Burke L.M., Gibala M.J., van Loon J.C. Nutritional strategies to promote postexercise recovery. Int J Sport Nutr Exerc Metab 2010:20(6);515-532. doi:10.1123/ijsnem.20.6.515
Berardi J.M., Noreen E.E., Lemon P.W.R. Recovery from a cycling time trial is enhanced with carbohydrate-protein supplementation vs. isoenergetic carbohydrate supplementation. J Intern Soc Sports Nutrition 2008;5:24. doi:10.1186/1550-2783-5-24
Betts J., Williams C., Duffy K., Gunner F. The influence of carbohydrate and protein ingestion during recovery from prolonged exercise on subsequent endurance performance. J Sports Sciences 2007;25(13):1449-1460. doi:10.1080/02640410701213459
Howarth K.R., Moreau N.A., Phillips S.M., and Gibala M.J. Coingestion of protein with carbohydrate during recovery from endurance exercise stimulates skeletal muscle protein synthesis in humans. J Appl Physiol 2009:106;1394–1402. doi:10.1152/japplphysiol.90333.2008
Jentjens R., Jeukendrup A. E. Determinants of post-exercise glycogen synthesis during short-term recovery. Sports Medicine 2003:33(2);117-144. doi:10.2165/00007256-200333020-00004
Millard-Stafford M., Childers W.L., Conger S.A., Kampfer A.J., Rahnert J.A. Recovery nutrition: timing and composition after endurance exercise. Curr Sports Med Rep 2008;7(4):193-201. doi:10.1249/JSR.0b013e31817fc0fd
Price T.B., Rothman D.L., Taylor R., Avison M.J., Shulman G.I., Shulman R.G. Human muscle glycogen resynthesis after exercise: insulin-dependent and –independent phases. J App Physiol 1994:76(1);104–111. doi:10.1152/jappl.1994.76.1.104
Schweitzer G.G., Kearney M.L., Mittendorfer B. Muscle glycogen: where did you come from, where did you go? J Physiol 2017;595(9):2771-2772. doi:10.1113/JP273536
van Loon L.J.C., Saris W.H.M., Kruijshoop M., Wagenmakers A.J.M. Maximizing postexercise muscle glycogen synthesis: carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures. Am J Clin Nutr 2000;72: 106-111. doi:10.1093/ajcn/72.1.106
Gli acidi grassi polinsaturi omega-6 sono i principali acidi grassi polinsaturi, detti anche PUFA, acronimo dell’inglese polyunsaturated fatty acids, nella dieta occidentale, circa il 90% dei tutti quelli presenti, essendo tra i costituenti della maggior parte dei grassi animali e vegetali.
All’interno della famiglia omega-6 uno degli acidi grassi più importanti e diffusi è l’acido linoleico, il precursore di tutti gli acidi grassi polinsaturi omega-6. È prodotto de novo dall’acido oleico, un acido grasso omega-9, solamente nelle piante in una reazione catalizzata dalla Δ12-desaturasi, dunque l’enzima che forma la famiglia degli acidi grassi polinsaturi omega-6 a partire da quella degli omega-9.
La Δ12-desaturasi catalizza l’inserimento di un doppio legame tra gli atomi di carbonio 6 e 7, numerati a partire dall’estremità metilica della molecola.
L’acido linoleico, insieme con con l’acido alfa-linolenico, è un prodotto primario della sintesi degli acidi grassi polinsaturi nelle piante.
Metabolismo degli Acidi Grassi Polinsaturi Omega-6
Gli animali, essendo privi della Δ12-desaturasi, non sono in grado di sintetizzare de novo ne l’acido linoleico ne tutta la famiglia degli omega-6 e sono obbligati ad assumerlo dagli alimenti vegetali e/o da animali che se ne cibino; per questo motivo gli omega-6 sono considerati acidi grassi essenziali, i cosiddetti EFA, acronimo dell’inglese essential fatty acids.
La loro essenzialità, in particolare proprio l’essenzialità dell’acido linoleico, è stata riportata per la prima volta nel 1929 da Burr e Burr.
Omega-6: da acido linoleico ad acido arachidonico
Gli animali sono in grado di allungare e desaturare l’acido linoleico ottenuto con la dieta in una cascata di reazioni che porta alla formazione di PUFA omega-6 a catena molto lunga.
L’acido linoleico viene prima desaturato ad acido gamma-linolenico, un altro importante acido grasso omega-6 con significativi effetti fisiologici, nella reazione catalizzata dalla Δ6-desaturasi. Si pensa che la velocità di questa reazione in certe condizioni, come negli anziani, in alcune situazioni patologiche e nei neonati prematuri, sia il fattore limitante della cascata; per questo motivo, e per il fatto che l’acido gamma-linolenico si trova in quantità relativamente piccole nella dieta, alcuni oli che lo contengono, quali gli oli di semi ottenuti da ribes, primula della sera e borragine, hanno attirato l’attenzione.
A sua volta l’acido gamma-linolenico può essere allungato ad acido diomo-gamma-linolenico in una reazione catalizzata da un elongasi. L’enzima catalizza l’aggiunta di due atomi di carbonio provenienti dal metabolismo del glucosio per allungare la catena dell’acido grasso, che può essere ulteriormente desaturato in quantità molto limitata a dare acido arachidonico, in una reazione catalizzata da un altro enzima limitante la velocità della cascata, la Δ5-desaturasi.
L’acido arachidonico può essere allungato e desaturato ad acido adrenico.
Va notato che gli acidi grassi polinsaturi della famiglia omega-6, e di ogni altra famiglia, possono essere interconvertiti attraverso processi enzimatici solo all’interno della stessa famiglia, non tra famiglie.
Gli acidi grassi polinsaturi con 20 atomi di carbonio appartenenti alle famiglie omega-6 e omega-3 sono i precursori degli eicosanoidi (prostaglandine, prostacicline, trombossani, e leucotrieni), potenti ormoni ad azione breve e locale.
Mentre la privazione di acidi grassi polinsaturi omega-3 causa la disfunzione in una vasta gamma di modalità comportamentali e fisiologiche, l’omissione dalla dieta degli acidi grassi polinsaturi omega-6 determina una palese disfunzione sistemica.
Negli oli di semi gli acidi grassi polinsaturi omega-6 con catena più lunga di 18 atomi di carbonio sono presenti solo in tracce mentre l’acido arachidonico si trova in tutti i tessuti animali e nei cibi di origine animale.
Bibliografia
Akoh C.C. and Min D.B. “Food lipids: chemistry, nutrition, and biotechnology” 3th ed. 2008
Aron H. Uber den Nahvert (On the nutritional value). Biochem Z. 1918;92:211–233 (German)
Bender D.A. “Benders’ dictionary of nutrition and food technology”. 2006, 8th Edition. Woodhead Publishing. Oxford
Bergstroem S., Danielsson H., Klenberg D. and Samuelsson B. The enzymatic conversion of essential fatty acids into prostaglandins. J Biol Chem 1964;239:PC4006-PC4008.
Burr G.O. and Burr M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. Nutr Rev 1973;31(8):148-149. doi:10.1111/j.1753-4887.1973.tb06008.x
Chow Ching K. “Fatty acids in foods and their health implication” 3th ed. 2008
Rosenthal M.D., Glew R.H. Mediacal biochemistry. Human metabolism in health and disease. John Wiley & Sons, Inc. 2009
Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3rd Edition. Elsevier health sciences, 2012
Van D., Beerthuis R.K., Nugteren D.H. and Vonkeman H. Enzymatic conversion of all-cis-polyunsaturated fatty acids into prostaglandins. Nature 1964;203:839-841
Gli acidi grassi polinsaturi omega-3 sono acidi grassi insaturi, dunque lipidi, che presentano un doppio legame in posizione 3 rispetto all’estremità metilica, detta anche estremità n, della catena carboniosa.[3]
Per gli esseri umani gli omega-3 più importanti sono:
l’acido alfa-linolenico o ALA o 18:3n-3, il più semplice, con 18 atomi di carbonio e tre doppi legami;
l’acido eicosapentaenoico o EPA o 20:5n-3, con 20 atomi di carbonio e 5 doppi legami;
l’acido docosaesaenoico o DHA o 22:6n-3, che con i suoi 22 atomi di carbonio e 6 doppi legami è il più complesso.
EPA e DHA sono definiti acidi grassi polinsaturi a catena lunga o LC-PUFA.
A causa della mancanza di due specifiche desaturasi, la delta-12 desaturasi (EC 1.14.19.6) e la delta-15 desaturasi (EC 1.14.19.13), che inseriscono doppi legami rispettivamente in posizione 6 e 3 dall’estremità metilica della catena carboniosa, i mammiferi non sono in grado di produrre l’acido linoleico o LA, capostipite della famiglia degli acidi grassi polinsaturi omega-6, e l’acido alfa-linolenico, a sua volta capostipite degli omega-3 e che deriva dall’acido linoleico.[9] L’acido linoleico e l’acido alfa-linolenico risultano pertanto essere acidi grassi essenziali.[34]
Molti animali, compreso l’uomo, hanno il corredo enzimatico necessario per sintetizzare, a partire dall’acido alfa-linolenico di origine alimentare, tutti gli altri omega-3, che quindi risultano essenziali solamente in assenza di ALA di origine alimentare, e per questo sono definiti acidi grassi semiessenziali.[9]
EPA e DHA sono componenti strutturali delle membrane cellulari che rappresentano la loro principale sede di deposito, in particolare nei tessuti muscolare e nervoso.[35] In ciò si differenziano dalla maggior parte degli altri acidi grassi che sono immagazzinati principalmente nei trigliceridi del tessuto adiposo.
Il DHA è il principale componente dei fosfolipidi delle membrane cellulari dei tessuti nervosi dei vertebrati, compresi i fotorecettori della retina, dove svolge importanti funzioni.[22][26][29][32] Oltre alla funzione strutturale, gli LC-PUFA omega-3 sono substrati per la produzione di mediatori lipidici bioattivi ad azione antiinfiammatoria quali eicosanoidi, maresine, resolvine, e protectine.[10][21]
Gli acidi grassi polinsaturi omega-3 sono essenziali nel corso dello sviluppo neurologico del feto, e la loro assunzione da parte della gestante è particolarmente importante nel terzo trimestre di gravidanza, fase in cui si verifica una crescita cerebrale significativa.[8] Nel corso della vita la loro assunzione è stata associata a una riduzione del rischio di sviluppare molte malattie croniche, in particolare le malattie cardiovascolari.[1][27][33]
Le fonti migliori per l’uomo sono i prodotti della pesca, specialmente quelli ottenuti da mari freddi.[3][9][36]
L’acido alfa-linolenico è prodotto dall’acido linoleico, un omega-6, nei plastidi del fitoplancton e delle piante terrestri vascolari, dove è presente la delta-15 desaturasi che inserisce un doppio legame tra gli atomi di carbonio 3 e 4, numerati a partire dall’estremità metilica, di LA.[3][9]
ALA va di seguito incontro a successive reazioni di desaturazione, catalizzate dalla delta-5 desaturasi (EC 1.14.19.44) e dalla delta-6 desaturasi (EC 1.14.19.3), e di allungamento, catalizzate dalla elongasi 5 (EC 2.3.1.199) e dalla elongasi 5 e/o dalla elongasi 2 (EC 2.3.1.199), e a una limitata beta-ossidazione nei perossisomi, a dare infine DHA.[6][8]
Va sottolineato che le piante terrestri vascolari non hanno la capacità di sintetizzare LC-PUFA omega-3, come EPA e DHA.[23]
Metabolismo degli Acidi Grassi Polinsaturi Omega-3
Gli enzimi che catalizzano la conversione dell’acido alfa-linolenico in DHA sono condivisi con le vie che portano alla sintesi degli acidi grassi polinsaturi omega-6, omega-7 e omega-9.[9]
Sembra che la delta-5 desaturasi e dalla delta-6 desaturasi abbiano come substrati preferenziali gli omega-3. Tuttavia, considerando che nella dieta occidentale il rapporto tra l’assunzione di ALA e LA è molto spostato a favore del secondo, ne risulterebbe favorita la via degli omega-6 rispetto alle altre.[38] Questa potrebbe essere una delle spiegazioni che giustificano il basso tasso di conversione dell’acido alfa-linolenico negli altri omega-3, sebbene anche la sintesi di acido arachidonico o ARA dall’acido linoleico sembra essere molto contenuta.
Da notare che sia la famiglia degli acidi grassi polinsaturi omega-3 che quella degli omega-6 inibiscono la formazione degli acidi grassi polinsaturi omega-9.
Biosintesi degli Omega-3 nell’uomo
L’uomo, così come molti altri animali, è in grado di allungare e desaturare l’acido alfa-linolenico sino a formare il DHA, via metabolica presente principalmente nel fegato e nel microcircolo cerebrale della barriera ematoencefalica, ma anche nell’endotelio celebrale e negli astrociti.[18] E’ tuttavia opinione comune che, al pari degli altri animali terrestri, abbia una limitata capacità di produrre gli LC-PUFA, e dunque necessiti di EPA e DHA di origine alimentare.[7]
E’ stato evidenziato che l’efficienza della sintesi diminuisce lungo la via: la conversione dell’acido alfa-linolenico a EPA è limitata, e il fattore limitante sarebbe l’attività della delta-6 desaturasi. La sintesi del acido docosaesaenoico sarebbe ancora più limitata della precedente.[7] Tuttavia recenti studi hanno dimostrato l’esistenza di un marcato polimorfismo globale nel cluster dei geni FADS, i geni che codificano per le desaturasi, in particolare per quelli che codificano per la delta-5 desaturasi e delta-6 desaturasi, rispettivamente FADS1 e FADS2, geni contigui presenti sul cromosoma 11q12.2.[6]
Dal confronto tra materiale genetico di soggetti vissuti nell’Età del Bronzo con quello di popolazioni contemporanee è stato possibile ottenere un quadro chiaro riguardo ai cambiamenti della frequenza allelica nel cluster dei geni FADS.[25] Differenze genotipiche che si sono riflesse in differenze fenotipiche.
Nelle popolazioni europee, il passaggio da una società di cacciatori-raccoglitori a una di agricoltori avrebbe determinato un aumento dell’assunzione di acido linoleico e acido alfa-linolenico e una riduzione dell’assunzione di EPA e ARA. La selezione avrebbe quindi favorito l’aplotipo associato all’aumento dell’espressione di FADS1 e alla diminuzione dell’espressione di FADS2.[6]
Questo è esattamente il pattern opposto a quello osservato negli Inuit, dove si ipotizza che la selezione abbia agito per favorire alleli che riducano la conversione dell’acido linoleico e dell’acido alfa-linolenico in LC-PUFA al fine di compensare il loro apporto relativamente alto con la dieta osservato in queste popolazioni.[13]
Gli altri animali hanno bisogno di EPA e DHA?
Gli organismi privi della delta-15 desaturasi non sono in grado di sintetizzare l’acido alfa-linolenico, e quindi tutta la famiglia degli omega-3, e, se necessario, sono obbligati a ottenerlo dai vegetali e/o da animali che se ne cibano. Ma non tutti gli animali hanno necessità di ottenere EPA e DHA dal cibo.
I vertebrati erbivori terrestri, come le mucche, soddisfano le loro necessità di LC-PUFA omega-3 producendoli a partire dall’acido alfa-linolenico ottenuto mangiando le parti verdi delle piante.[19]
E ci sono animali che non necessitano di EPA e DHA, e ne sono praticamente privi. Tra questi gli insetti terrestri, nei quali si ritrovano solo bassissimi livelli di EPA, paragonabili a quelli delle vitamine. In essi l’EPA è sintetizzato a partire dall’ALA di origine alimentare e utilizzato per la produzione di eicosanoidi.[16]
Invece, gli insetti acquatici presentano elevati livelli di EPA, mentre il DHA è praticamente assente.[31]
Sono molto ricchi in EPA e DHA diverse classi di fitoplancton come le Cryptophyceae, le Dinophyceae, mentre sono molto ricche in EPA le Bacillariophyceae o diatomee. In generale le microalghe rappresentano i produttori primari di EPA e DHA. Gli ecosistemi acquatici sono quindi la principale fonte di LC-PUFA omega-3 nella biosfera.[2][28] EPA e DHA sintetizzati in queste alghe unicellulari sono trasferiti lungo la catena alimentare, passando dagli invertebrati ai pesci è poi agli animali terrestri, esseri umani inclusi. Dunque, dalle microalghe agli esseri umani.[16]
Benefici
Gli acidi grassi polinsaturi omega-3 sono componenti essenziali di una dieta salutare e bilanciata in quanto necessari nel corso dello sviluppo, a partire dalla vita fetale, e associati, nel corso della vita, a miglioramenti della salute e alla riduzione del rischio di malattia.[8][26][36]
Studi epidemiologici hanno ad esempio associato elevati livelli di assunzione di EPA e DHA con tassi di mortalità per malattie cardiovascolari, specialmente cardiache, più bassi di quelli attesi in soggetti sani, probabile conseguenza del miglioramento di molti fattori di rischio, in particolare dei livelli plasmatici dei trigliceridi, del colesterolo HDL e della proteina C-reattiva, e miglioramenti dei valori della pressione, sia sistolica che diastoliche, e della frequenza cardiaca.[1][27]
EPA e DHA si sono dimostrati utili anche nel trattamento di malattie quali l’artrite reumatoide, e potrebbero essere utili nel trattamento di altre condizioni infiammatorie quali l’asma, la psoriasi o le malattie infiammatorie croniche intestinali, grazie alla loro capacità di modulare molti aspetti dei processi infiammatori.[33]
Al contrario, gli LC-PUFA omega-3 sembrano avere effetti minimi o nulli sulla prevenzione e il trattamento del diabete mellito di tipo 2.[5]
Rapporto Omega-3/Omega-6
Molte prove epidemiologiche suggeriscono che il consumo di una dieta con un basso valore del rapporto omega-3/omega-6 abbia influenzato negativamente la salute umana, concorrendo alla promozione, probabilmente assieme a fattori quali il fumo e la sedentarietà, dello sviluppo delle principali classi di patologie.[8]
Sono stati infatti osservati tassi più bassi di incidenza di cancro, autoimmunità e malattia coronarica nelle popolazioni che consumano una dieta con un alto valore del rapporto omega-3/omega-6, in particolare negli Eschimesi, Giapponesi e in altre società dove è elevato il consumo di pesce.[20][37]
Nonostante ciò, la dieta di tipo occidentale è divenuta via via più ricca in acidi grassi saturi, povera in acidi grassi polinsaturi omega-3 e ricca in acidi grassi polinsaturi omega-6, con una rapporto omega-3/omega-6 compreso tra 1/10 e 1/20, quindi molto lontano dal rapporto raccomandato di 1:5.[33]
Il basso valore del rapporto omega-3/omega-6 è dovuto a diversi fattori, alcuni dei quali sono di seguito elencati.
Sebbene gli alimenti vegetali selvatici siano in genere ricchi in omega-3, nell’agricoltura industriale le colture ricche in omega-6 hanno avuto molto più successo di quelle ricche in omega-3.[3]
Il basso consumo di prodotti della pesca e di olio di pesce.[33]
L’elevato consumo di animali allevati con mangimi a base di mais, quali polli, bovini e suini.
A questo va aggiunto anche il fatto che il contenuto in omega-3 di alcune specie di pesci allevate è più basso rispetto a quello delle controparti selvatiche.[9]
L’elevato consumo di oli ricchi in omega-6 e poveri in omega-3, come l’olio di cartamo, girasole, di semi di soia, e di mais.[4]
Nota: il rapporto omega-3/omega-6 non sembra avere influenza ne sulla prevenzione ne sul trattamento del diabete di tipo 2.[5]
Effetti a livello molecolare di EPA e DHA
Negli ultimi anni si stanno chiarendo i meccanismi molecolari alla base degli effetti funzionali attribuiti agli acidi grassi polinsaturi omega-3, in particolare a EPA e DHA, e la maggior parte di questi richiedono l’incorporazione dei due acidi grassi nei fosfolipidi di membrana.[23]
Gli omega-3 sono componenti strutturali delle membrane cellulari, dove svolgono un ruolo essenziale nel regolarne la fluidità.[35] Grazie a questo effetto gli omega-3, in particolare EPA e DHA, sono in grado di modulare le risposte cellulari dipendenti da proteine di membrana. Questo si è rivelato essere particolarmente importante nell’occhio, dove la presenza del DHA permette un’attività ottimale della rodopsina.[29] L’azione sulla fluidità di membrana è poi essenziale per gli animali che vivono in acque fredde, nei quali EPA e DHA svolgono anche una funzione di antigelo naturali.
EPA e DHA sono in grado di modificare la formazione delle zattere lipidiche, microdomini con una composizione lipidica specifica che possono agire come piattaforme per l’azione dei recettori e per l’inizio di vie di segnalazione intracellulare. Modificandone la formazione influenzano le vie di segnalazione intracellulare in tipi cellulari differenti, come i neuroni, le cellule del sistema immunitario e le cellule cancerose. In questo modo EPA e DHA sono in grado di modulare l’attivazione di fattori di trascrizione, come NF-κB, PPARs e SREBPs, e di conseguenza i pattern di espressione genica da essi regolati. Questo ruolo è centrale rispetto alla loro funzione di controllo del metabolismo degli acidi grassi e dei trigliceridi, dell’infiammazione e della differenziazione adipocitaria.[8]
ARA, EPA, e DHA sono substrati per la produzione di numerosi mediatori lipidici, tra cui spiccano gli eicosanoidi, molecole con un ruolo ben definito nella regolazione dell’infiammazione, immunità, aggregazione piastrinica, funzione renale e contrazione della muscolatura liscia.[21]
Gli eicosanoidi prodotti dall’acido arachidonico, che, dal punto di vista quantitativo, è il maggior substrato per la loro sintesi, hanno importanti ruoli fisiologici, ma un loro eccesso è stato associato a numerosi processi patologici.[8] L’aumento del contenuto di EPA e DHA nei fosfolipidi di membrana si accompagna a una riduzione del contenuto di ARA, a una riduzione nella produzione di mediatori lipidici derivanti da ARA, e a un aumento della produzione di mediatori lipidici derivanti dai due omega-3. In aggiunta, tra le molecole derivanti da EPA e DHA ci sono alcuni eicosanoidi analoghi a quelli derivati dall’acido arachidonico, sebbene dotati di minor attività biologica, le resolvine, e, per quanto riguarda DHA, le protectine e le maresine. Queste molecole sembrano essere responsabili di molte delle azioni antiinfiammatorie immuno-modulatrici attribuite ai due acidi grassi polinsaturi omega-3.[10]
E’ stato dimostrato che EPA e DHA possono svolgere un ruolo in forma libera. Sono infatti in grado di agire direttamente attraverso recettori di membrana accoppiati alle proteine G, modulandone l’attività o legandosi a specifici recettori nucleari, come il DHA su PPARα.[30]
Infine, riducono l’assorbimento degli acidi grassi polinsaturi omega-6, e, a livello enzimatico inibiscono competitivamente la ciclo ossigenasi e la lipossigenasi, e competono per le aciltransferasi.[3]
Le fonti più importanti di EPA e DHA per l’uomo
In generale i pesci e gli invertebrati acquatici, come i crostacei e i molluschi, sono la fonte più importante di EPA e DHA per l’uomo.[3][9][36] Questi animali sono in grado di convertire l’acido alfa-linolenico in EPA e DHA oltre che di assumerlo con il cibo, quindi, in ultima analisi dal fitoplancton.[16] Inoltre, considerando il solo DHA, questi è presente in concentrazione elevata in molti oli di pesce, specialmente in quelli di pesci cresciuti in acque fredde. Si noti però che molti di questi oli sono ricchi anche di acidi grassi saturi.[24]
Per chi non mangia prodotti della pesca, il fegato degli animali terrestri ma anche la selvaggina dell’ordine dei Passeriformi rappresentano una buona fonte di EPA e DHA.[14][15]
Riguardo gli apporti giornalieri non è ancora chiaro quale siano i valori raccomandati. Nella tabella seguente sono riportati i valori suggeriti dalla FAO delle Nazioni Unite e dalla autorità europea per la sicurezza alimentare (EFSA).[11][12]
Categoria
Raccomandazioni
FAO
Uomini adulti e donne adulte non gravide e non in allattamento
Almeno 250 mg di EPA + DHA al giorno
FAO
Donne gravide o in allattamento
Almeno 300 mg di EPA + DHA al giorno di cui almeno 200 mg dovrebbero essere di DHA
FAO
Bambini di età compresa tra i 2 e i 4 anni
100-150 mg di EPA + DHA al giorno
FAO
Bambini dai 4 ai 6 anni
150-200 mg di EPA + DHA al giorno
FAO
Bambini dai 6 ai 10 anni
200-250 mg di EPA + DHA al giorno
EFSA
Uomini adulti e donne adulte non gravide
L’assunzione adeguata è di 250 mg di EPA + DHA al giorno
EFSA
Donne gravide
Ulteriori 100-200 mg di DHA al giorno
EFSA
Neonati e bambini da 6 mesi a 2 anni
100 mg di DHA al giorno
EFSA
Bambini dai 2 ai 18 anni
“Coerente con gli adulti”
Omega-3 negli alimenti
Gli acidi grassi polinsaturi sono particolarmente sensibili all’ossidazione a seguito del riscaldamento. I vari processi cui è sottoposto il cibo nella fase di preparazione potrebbero quindi ridurne il contenuto.[3] Tuttavia questo è vero solo in parte. Negli alimenti i due acidi grassi polinsaturi omega-3 non sono presenti in forma chimica pura, ma per la maggior parte esterificati nei fosfolipidi di membrana, e, in questa forma, risultano molto meno suscettibili all’ossidazione.[9]
Quando si considera il contenuto in EPA e DHA negli alimenti, esprimerlo come percentuale degli acidi grassi totali anziché come contenuto assoluto, mg/g di peso umido, induce in un errore. Se ad esempio si considera un pesce grasso come il salmone, il suo contenuto assoluto di EPA + DHA è alto, circa 8 mg/g di peso umido, mentre se si esprime in percentuale si ottiene il 20%. Di contro il merluzzo atlantico presenta una percentuale di EPA + DHA di oltre il 40%, ma un contenuto assoluto inferiore a 3 mg/g di peso umido. In questo caso l’alta percentuale è dovuta al fatto che il pesce in esame è magro, mentre nei pesci grassi il contenuto in EPA + DHA è “diluito” dalle elevate riserve di acidi grassi presenti nel tessuto adiposo dell’animale.[17]
Se invece si esprime il contenuto di EPA + DHA in mg/g di prodotto, non si osserva alcuna riduzione a seguito della maggior parte dei trattamenti cui il cibo è sottoposto nella fase di preparazione.[16]
Bibliografia
^ ab AbuMweis S., Jew S., Tayyem R. Agraib L. Eicosapentaenoic acid and docosahexaenoic acid containing supplements modulate risk factors for cardiovascular disease: a meta-analysis of randomised placebo-control human clinical. J Hum Nutr Diet 2018;31(1):67-84. doi:10.1111/jhn.12493
^ Ahlgren G., Lundstedt L., Brett M., Forsberg C. Lipid composition and food quality of some freshwater phytoplankton for cladoceran zooplankters. J Plankton Res 1990;12(4):809-18. doi:10.1093/plankt/12.4.809
^ abcdefg Akoh C.C. and Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 3th ed. 2008
^ Blasbalg T.L., Hibbeln J.R., Ramsden C.E., Majchrzak S..F, Rawlings R.R. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr 2011;93(5):950-62. doi:10.3945/ajcn.110.006643
^ ab Brown T.J., Brainard J., Song F., Wang X., Abdelhamid A., Hooper L. Omega-3, omega-6, and total dietary polyunsaturated fat for prevention and treatment of type 2 diabetes mellitus: systematic review and meta-analysis of randomised controlled trials. BMJ 2019;366:l4697. doi:10.1136/bmj.l4697
^ abc Buckley M.T., Racimo F., Allentoft M.E., et al. Selection in Europeans on fatty acid desaturases associated with dietary changes. Mol Biol Evol 2017;34(6):1307-1318. doi:10.1093/molbev/msx103
^ ab Burdge G.C., Wootton S.A. Conversion of α-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br J Nutr 2002;88(4):411-420. doi:10.1079/BJN2002689
^ abcdef Calder P.C. Very long-chain n-3 fatty acids and human health: fact, fiction and the future. Proc Nutr Soc 2018 77(1):52-72. doi:10.1017/S0029665117003950
^ abcdefgh Chow Ching K. Fatty acids in foods and their health implication. 3th ed. 2008
^ ab Duvall M.G., Levy B.D. DHA- and EPA-derived resolvins, protectins, and maresins in airway inflammation. Eur J Pharmacol 2016;785:144-155. doi:10.1016/j.ejphar.2015.11.001
^ EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific opinion on dietary reference values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. 2010. doi:10.2903/j.efsa.2010.1461
^ FAO. Global recommendations for EPA and DHA intake (As of 30 June 2014)
^ Fumagalli M., Moltke I., Grarup N., Racimo F., Bjerregaard P., Jørgensen M.E., Korneliussen T.S., Gerbault P., Skotte L., Linneberg A., Christensen C., Brandslund I., Jørgensen T., Huerta-Sánchez E., Schmidt E.B., Pedersen O., Hansen T., Albrechtsen A., Nielsen R.. Greenlandic Inuit show genetic signatures of diet and climate adaptation. Science 2015;349(6254):1343-7. doi:10.1126/science.aab2319
^ Gladyshev M.I., Makhutova O.N., Gubanenko G.A.; Rechkina E.A., Kalachova G.S., Sushchik N.N. Livers of terrestrial production animals as a source of long-chain polyunsaturated fatty acids for humans: an alternative to fish? Eur J Lipid Sci Technol 2015;117:1417-1421. doi:10.1002/ejlt.201400449
^ Gladyshev M.I., Popova O.N., Makhutova, O.N., Zinchenko, T.D., Golovatyuk, L.V., Yurchenko, Y.A., Kalachova, G.S., Krylov, A.V., Sushchik, N.N. Comparison of fatty acid compositions in birds feeding in aquatic and terrestrial ecosystems. Contemp Probl Ecol 2016;9:503-513. doi:10.1134/S1995425516040065
^ abcd Gladyshev M.I. and Sushchik N.N. Long-chain omega-3 polyunsaturated fatty acids in natural ecosystems and the human diet: assumptions and challenges. Biomolecules 2019;9(9):485. doi:10.3390/biom9090485
^ Gladyshev M.I., Sushchik N.N., Tolomeev A.P., Dgebuadze Y.Y. Meta-analysis of factors associated with omega-3 fatty acid contents of wild fish. Rev Fish Biol Fish 2018;28,:277-299. doi:10.1007/s11160-017-9511-0
^ Kim H-Y. Novel metabolism of docosahexaenoic acid in neural cells. J Biol Chem 2007;282(26):18661-18665. doi:10.1074/jbc.R700015200
^ Kouba M., Mourot J. A review of nutritional effects on fat composition of animal products with special emphasis on n-3 polyunsaturated fatty acids. Biochimie 2011;93(1):13-7. doi:10.1016/j.biochi.2010.02.027
^ Kromann N., Green A. Epidemiological studies in the Upernavik district, Greenland. Incidence of some chronic diseases 1950-1974. Acta Med Scand 1980;208(5):401-6. doi:10.1111/j.0954-6820.1980.tb01221.x
^ Lauritzen L., Hansen H.S., Jørgensen M.H., Michaelsen K.F. The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Prog Lipid Res 2001;40(1-2):1-94. doi:10.1016/s0163-7827(00)00017-5
^ ab Lee J.M., Lee H., Kang S. and Park W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016;8(1):23. doi:10.3390/nu8010023
^ Mason R.P., Sherratt S.C.R. Omega-3 fatty acid fish oil dietary supplements contain saturated fats and oxidized lipids that may interfere with their intended biological benefits. Biochem Biophys Res Commun 2017;483(1):425-429. doi:10.1016/j.bbrc.2016.12.127
^ Mathieson I., Lazaridis I., Rohland N., Mallick S., Patterson N., Roodenberg S.A., Harney E., Stewardson K., Fernandes D., Novak M., et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature 2015;528:499-503. doi:10.1038/nature16152
^ ab McCann J.C., Ames B.N. Is docosahexaenoic acid, an n-3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am J Clin Nutr 2005;82(2):281-95. doi:10.1093/ajcn.82.2.281
^ ab Mensink R.P., Zock P.L., Kester A.D., Katan M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: a meta-analysis of 60 controlled trials. Am J Clin Nutr 2003;77(5):1146-55. doi:10.1093/ajcn/77.5.1146
^ Muller-Navarra D.C. Biochemical versus mineral limitation in Daphnia. Limnol Oceanogr 1995;40:1209-1214. doi:10.4319/lo.1995.40.7.1209
^ ab Niu S.L., Mitchell D.C., Lim S.Y., Wen Z.M., Kim H.Y., Salem N. Jr, Litman B.J. Reduced G protein-coupled signaling efficiency in retinal rod outer segments in response to n-3 fatty acid deficiency. J Biol Chem 2004;279(30):31098-104. doi:10.1074/jbc.M404376200
^ Oh D.Y., Talukdar S., Bae E.J., Imamura T., Morinaga H., Fan WQ, Li P., Lu W.J., Watkins S.M., Olefsky J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010 142(5):687-698. doi:10.1016/j.cell.2010.07.041
^ Popova O.N., Haritonov A.Y., Sushchik N.N., Makhutova O.N., Kalachova G.S., Kolmakova A.A., Gladyshev M.I. Export of aquatic productivity, including highly unsaturated fatty acids, to terrestrial ecosystems via Odonata. Sci Total Environ 2017;581-582:40-48. doi:10.1016/j.scitotenv.2017.01.017
^ Salem N. Jr, Litman B., Kim H.Y., Gawrisch K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001;36(9):945-59. doi:10.1007/s11745-001-0805-6
^ abcd Simopoulos A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp Biol Med 2008;233(6):6746-88. doi:10.1016/S0753-3322(02)00253-6
^ Smith W., Mukhopadhyay R. Essential fatty acids: the work of George and Mildred Burr. J Biol Chem 2012;287(42):35439-35441. doi:10.1074/jbc.O112.000005
^ ab Stubbs C.D., Smith A.D. The modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochim Biophys Acta 1984;779(1):89-137. doi:10.1016/0304-4157(84)90005-4
^ abc Tocher D.R., Betancor M.B., Sprague M., Olsen R.E., Napier J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: bridging the gap between supply and demand. Nutrients 2019;11(1):89. doi:10.3390/nu11010089
^ Yano K., MacLean C.J., Reed D.M., Shimizu Y., Sasaki H., Kodama K., Kato H., Kagan A. A comparison of the 12-year mortality and predictive factors of coronary heart disease among Japanese men in Japan and Hawaii. Am J Epidemiol 1988;127(3):476-87. doi:10.1093/oxfordjournals.aje.a114824
^ Williams C.M., Burdge G. Long-chain n-3 PUFA: plant v. marine sources. Proc Nutr Soc 2006;65(1):42-50. doi:10.1079/pns2005473
Gli acidi grassi essenziali o EFA sono acidi grassi insaturi che non possono essere sintetizzati dagli animali e, al pari degli altri nutrienti essenziali, devono essere ottenuti con la dieta. Nello specifico sono l’acido linoleico e l’acido alfa-linolenico.
L’incapacità degli animali di sintetizzare i due acidi grassi deriva dalla mancanza di due desaturasi, la delta-12 desaturasi (EC 1.14.19.6) e la delta-15 desaturasi (EC 1.14.19.25), enzimi in grado di inserire doppi legami cis oltre il carbonio 9.[1][9][15] Le due desaturasi sono presenti in organismi vegetali e microrganismi quali batteri, funghi e muffe.[10]
La delta-12 desaturasi catalizza la sintesi dell’acido linoleico a partire dall’acido oleico inserendo un doppio legame sul carbonio 12, quindi tra i carboni 6 e 7 se numerati dall’estremità metilica della molecola.[15]
La delta-15 desaturasi catalizza la sintesi dell’acido alfa-linolenico a partire dall’acido linoleico, inserendo un doppio legame sul carbonio 15, quindi tra i carboni 3 e 4 dall’estremità metilica della molecola. L’enzima è presente solo nel reticolo endoplasmatico e nei plastidi del fitoplancton e delle piante terrestri vascolari.[1][9]
Biosintesi degli Acidi Grassi Essenziali
L’acido linoleico e l’acido alfa-linolenico sono i capostipiti rispettivamente delle famiglie degli acidi grassi polinsaturi omega-6 e degli acidi grassi polinsaturi omega-3. Nel caso di mancanza nella dieta dei due acidi grassi essenziali, una condizione piuttosto rara, anche gli altri acidi grassi omega-6 e omega-3 diventano essenziali; per questo motivo questi lipidi da alcuni sono definiti acidi grassi semiessenziali.[9]
Nota: gli acidi grassi essenziali sono acidi grassi polinsaturi, ma non tutti gli acidi grassi polinsaturi sono essenziali, come quelli appartenenti alle famiglie degli omega-7 e omega-9.
La prima indicazione dell’esistenza degli acidi grassi essenziali risale al 1918, quando Hans Aron suggerì che il grasso assunto con la dieta potesse essere essenziale per il normale accrescimento degli animali e che, oltre al suo contributo calorico, ci fosse uno speciale valore nutritivo intrinseco derivante dalla presenza di alcune specifiche molecole lipidiche.[2]
Nel 1927, Herbert M. Evans e George Oswald Burr dimostrarono che, nonostante l’aggiunta alla dieta delle vitamine liposolubili A, D ed E, una carenza di grasso influenzava in modo grave sia la crescita che la riproduzione degli animali da esperimento, suggerendo la presenza in esso di una nuova sostanza indispensabile, che chiamarono vitamina F.[11]
Nel 1929, undici anni dopo il lavoro di Aron, Burr e sua moglie Mildred Lawson ipotizzarono che gli animali a sangue caldo non fossero in grado di sintetizzare quantità apprezzabili di certi acidi grassi e, un anno più tardi, scoprirono che l’acido linoleico era essenziale per gli animali.[7] E’ a questi due ricercatori che si deve la locuzione “acido grasso essenziale”.[8][19]
E fu solamente nel 1958, grazie al lavoro di Arild Hansen, che si ebbe la prima descrizione di una deficienza di acidi grassi essenziali nell’uomo, in particolare in neonati alimentati con una dieta a base di latte in cui tali acidi grassi erano assenti.[14]
Nel 1964, grazie alle ricerche di Van Dorp e Bergstroem, fu scoperta una delle loro funzioni biologiche, quella di essere precursori per la sintesi delle prostaglandine.[3][21]
Funzioni
Gli acidi grassi essenziali svolgono importanti funzioni biologiche.
L’acido linoleico e l’acido alfa-linolenico possono essere utilizzati a scopi energetici, quindi andare incontro a beta-ossidazione.[22]
A partire da essi gli animali, compreso l’uomo, sono in grado di sintetizzare, sebbene con efficienza variabile, gli altri membri delle famiglie degli omega-6, ad esempio l’acido arachidonico, e omega-3, ad esempio l’acido eicosapentaenoico e l’acido docosaesaenoico.[5][6][18]
Gli acidi grassi essenziali sono componenti strutturali delle membrane cellulari, di cui, tra le altre cose, ne modulano la fluidità.[20][22]
Sono essenziali, in particolare l’acido linoleico presente negli sfingolipidi dello strato corneo della pelle, per la formazione della barriera che si oppone alla perdita d’acqua dalla pelle stessa.[23]
Hanno un ruolo cruciale nella prevenzione di molte malattie, in particolare delle malattie coronariche.[12][16]
Fonti alimentari
L’acido linoleico è il più abbondante acido grasso polinsaturo nella dieta occidentale, e rappresenta l’85-90% di tutti gli omega-6 ingeriti.[13] Le principali fonti per l’uomo sono diversi oli vegetali e i semi di molte piante.[4]
Alimenti
Acido linoleico (mg/g)
Olio di cartamo
∼ 740
Olio di girasole
∼ 600
Olio di soia
∼ 530
Olio di mais
∼ 500
Olio di semi di cotone
∼ 480
Noci
∼ 340
Noci brasiliane
∼ 250
Olio di arachidi
∼ 240
Olio di colza
∼ 190
Arachidi
∼ 140
Olio di semi di lino
∼ 135
L’acido linoleico è presente in discreta quantità anche in prodotti di origine animale, come le uova di gallina o il lardo, ma solo perché gli animali sono stati allevati con mangimi che lo contengono.[9]
Da notare che alcune tra le principali fonti di acido linoleico, come le noci, l’olio di semi di lino, l’olio di soia, e l’olio di colza sono anche ricche di acido alfa-linolenico.[17]
Tra le fonti più ricche di acido alfa-linolenico presenti nell’alimentazione umana si ritrovano l’olio di semi di lino, circa 550 mg/g, l’olio di colza, circa 85 mg/g, e l’olio di soia, circa 75 mg/g.
Altri alimenti ricchi di acido alfa-linolenico sono le noci, con un contenuto di circa 70 mg/g e i semi di soia, circa 10 mg/g.[1]
Bibliografia
^ abc Akoh C.C. and Min D.B. Food lipids: chemistry, nutrition, and biotechnology. 3th Edition. CRC Press, Taylor & Francis Group, 2008
^ Aron H. Uber den Nahrwert. Biochem Z 1918; 92: 211-33
^ Bergstroem S., Danielsson H., Klenberg D., Samuelsson B. The enzymatic conversion of essential fatty acids into prostaglandins. J Biol Chem 1964;239:PC4006-8. doi:10.1016/S0021-9258(18)91234-2
^ Blasbalg T. L., Hibbeln J. R., Ramsden C. E., Majchrzak S. F. & Rawlings R. R. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr 2011;93(5):950-962. doi:10.3945/ajcn.110.006643
^ Burdge G.C, Jones A.E, Wootton S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of α-linolenic acid metabolism in young men. Br J Nutr 2002;88(4):355-364. doi:10.1079/BJN2002662
^ Burdge G.C., Wootton S.A. Conversion of α-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br J Nutr 2002;88(4):411-420. doi:10.1079/BJN2002689
^ Burr G. and Burr M. A new deficiency disease produced by the rigid exclusion of fat from the diet. J Biol Chem 1929;82:345-367. doi:10.1016/S0021-9258(20)78281-5
^ Burr G.O., Burr M.M., Miller E.S. On the fatty acids essential in nutrition. III. J Biol Chem 1932;97(1):1-9. doi:10.1016/S0021-9258(18)76213-3
^ abcd Chow Ching K. Fatty acids in foods and their health implication. 3th Edition. CRC Press, Taylor & Francis Group, 2008
^ Das U.N. Essential fatty acids: biochemistry, physiology and pathology. Biotechnol J 2006;1;420-439. doi:10.1002/biot.200600012
^ Evans H. M. and G. O. Burr. A new dietary deficiency with highly purified diets. III. The beneficial effect of fat in the diet. Proc Soc Exp Biol Med 1928;25:390-397. doi:10.3181/00379727-25-3867
^ Farvid M.S., Ding M., Pan A., Sun Q., Chiuve S.E., Steffen L.M., Willett W.C., Hu F.B. Dietary linoleic acid and risk of coronary heart disease: a systematic review and meta-analysis of prospective cohort studies. Circulation 2014;130:1568-1578. doi:10.1161/CIRCULATIONAHA.114.010236
^ Guyenet S.J., Carlson S.E. Increase in adipose tissue linoleic acid of US adults in the last half century. Adv Nutr 2015;6(6):660-664. doi:https://DOI.org/10.3945/an.115.009944
^ Hansen A.E., Haggard M.E., Boelsche A.N., Adam D.J., Wiese H.F. Essential fatty acids in infant nutrition. III. Clinical manifestations of linoleic acid deficiency. J Nutr 1958;66(4):565-76. doi:10.1093/jn/66.4.565
^ ab Malcicka M., Visser B., Ellers J. An evolutionary perspective on linoleic acid synthesis in animals. Evol Biol 2018;45:15-26. doi:10.1007/s11692-017-9436-5
^ Mensink R.P., Zock P.L., Kester A.D., Katan M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: a meta-analysis of 60 controlled trials. Am J Clin Nutr 2003;77(5):1146-55. doi:10.1093/ajcn/77.5.1146
^ Rajaram S. Health benefits of plant-derived α-linolenic acid. Am J Clin Nutr 2014;100 Suppl 1:443S-8S. doi:10.3945/ajcn.113.071514
^ Rett B.S., Whelan J. Increasing dietary linoleic acid does not increase tissue arachidonic acid content in adults consuming Western-type diets: a systematic review. Nutr Metab (Lond) 2011;8:36. doi:10.1186/1743-7075-8-36
^ Smith W., Mukhopadhyay R. Essential fatty acids: the work of George and Mildred Burr. J Biol Chem 2012;287(42):35439-35441. doi:10.1074/jbc.O112.000005
^ Stubbs C.D., Smith A.D. The modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochim Biophys Acta 1984;779(1):89-137. doi:10.1016/0304-4157(84)90005-4
^ Van Dorp D.A., Beerthuis R.K., Nugteren D.H. and Vonkeman H. Enzymatic conversion of all-cis-polyunsaturated fatty acids into prostaglandins. Nature 1964;203:839-841. doi:10.1038/203839a0
^ ab Whelan J. and Fritsche K. Linoleic acid. Adv Nutr 2013;4(3): 311-312. doi:10.3945/an.113.003772
^ Ziboh V.A. Prostaglandins, leukotrienes, and hydroxy fatty acids in epidermis. Semin Dermatol 1992;11(2):114-20
Lo zucchero invertito corrisponde a saccarosio parzialmente o totalmente scisso in fruttosio e glucosio e, a prescindere dal procedimento utilizzato, la soluzione ottenuta contiene una eguale quantità dei due carboidrati.
Inoltre, a seconda del prodotto, può essere presente anche saccarosio non scisso.
Preparazione dello zucchero invertito
La rottura della molecola del saccarosio può avvenire in una reazione catalizzata da enzimi, quali:
la saccarasi-isomaltasi (EC 3.2.1.48), attiva a livello del nostro intestino dove è coinvolta nella digestione dei carboidrati;
la invertasi, enzima secreto dalle api nel miele e utilizzato a livello industriale per ottenere lo zucchero invertito.
Un altro procedimento ricorre all’azione di acidi, come avviene in parte nel nostro stomaco e come avveniva in passato, e ancora avviene, a livello casalingo e industriale. Venivano utilizzati acido solforico o cloridrico, scaldando con cautela per un certo periodo la soluzione; la reazione infatti è tanto più veloce quanto più è acida la soluzione a prescindere dal tipo di acido utilizzato, e maggiore è la temperatura. L’acidità veniva poi tamponata con sostanze alcaline quali la soda o il bicarbonato di sodio.
Un procedimento chimicamente simile a quello appena descritto si verifica quando si preparano cibi acidi. Un esempio è la preparazione di marmellate e confetture di frutta dove sono realizzate entrambe le condizioni di acidità, naturalmente, e alte temperature, attraverso il riscaldamento. Situazione analoga quando si dolcificano i succhi di frutta con saccarosio.
La reazione procede anche a temperatura ambiente, ovviamente più lentamente.
Questo che significa dal punto di vista pratico?
Significa che anche dolci e preparazioni acide, anche quelle già viste, durante la conservazione vanno incontro a una lenta reazione di inversione del saccarosio contenuto/residuo, con conseguente modificazione della dolcezza, poiché lo zucchero invertito alle basse temperature è più dolce, grazie alla presenza del fruttosio, e assunzione di un profilo gustativo differente.
Utilizzi
E’ utilizzato soprattutto in pasticceria e nella produzione del gelato grazie ad alcune sue peculiari caratteristiche.
Ha una maggiore affinità per l’acqua rispetto al saccarosio, per cui mantiene più umidi i prodotti in cui si trova: ad esempio, le torte fatte con lo zucchero invertito si seccano meno facilmente.
Evita o ritarda la formazione di cristalli, in quanto i due monosaccaridi formano meno cristalli rispetto al saccarosio, proprietà utile in pasticceria per glasse o coperture.
Ha un punto di congelamento più basso.
Incrementa, anche se di poco, la dolcezza del prodotto in cui è stato aggiunto poiché è più dolce di una eguale quantità di saccarosio. Va comunque ricordato che la dolcezza del fruttosio dipende dalla temperatura a cui si trova.
Può partecipare alla reazione di Maillard, cosa che il saccarosio non può fare, contribuendo così al colore e al gusto di alcuni prodotti da forno.
Da notare che il miele, essendo privo di saccarosio, ha quasi la stessa composizione in fruttosio e glucosio dello zucchero invertito al 100% (il fruttosio è leggermente più abbondante del glucosio). Quindi un miele, magari non molto aromatico, diluito in acqua può fare le veci di uno prodotto ottenuto industriale.
Bibliografia
Belitz .H.-D., Grosch W., Schieberle P. “Food Chemistry” 4th ed. Springer, 2009
Bender D.A. “Benders’ Dictionary of Nutrition and Food Technology”. 8th Edition. Woodhead Publishing. Oxford, 2006
Jordan S. Commercial invert sugar. Ind Eng Chem 1924;16(3):307-310. doi:10.1021/ie50171a037
Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3rd Edition. Elsevier health sciences, 2012
Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3rd Edition. Elsevier health sciences, 2013 [Google eBooks]
Biochemistry and metabolism
Utilizziamo i cookie per essere sicuri che tu possa avere la migliore esperienza sul nostro sito. Se continui ad utilizzare questo sito noi assumiamo che tu ne sia felice.OkPrivacy policy

 Questi
Questi  Rappresentando la principale fonte di lipidi, quando associato a un basso consumo di prodotti di origine animale ad alto contenuto di grassi, assicura un elevato rapporto tra acidi grassi monoinsaturi e
Rappresentando la principale fonte di lipidi, quando associato a un basso consumo di prodotti di origine animale ad alto contenuto di grassi, assicura un elevato rapporto tra acidi grassi monoinsaturi e 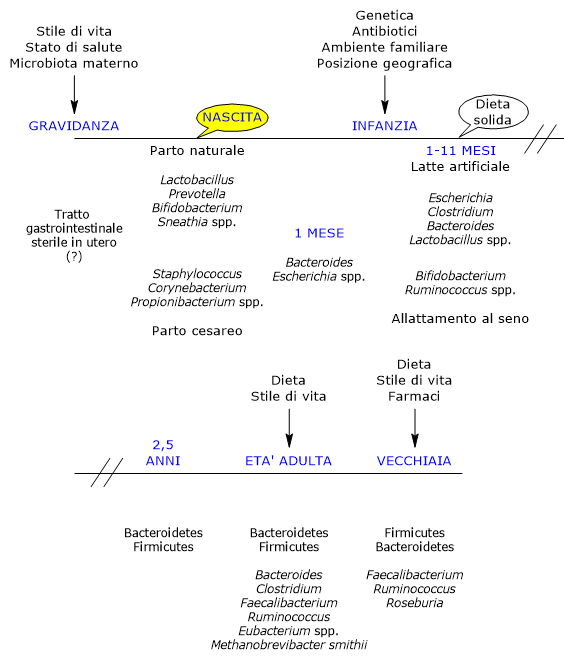
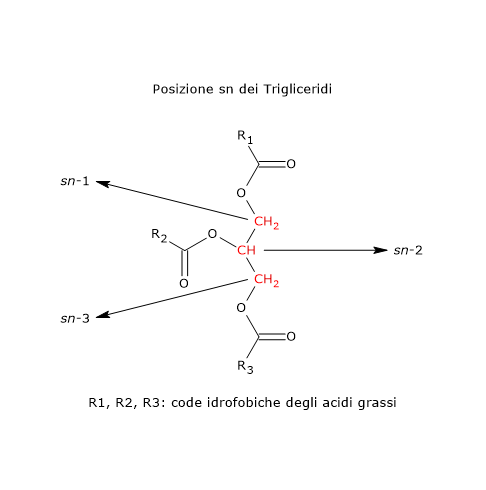
 Altre molecole presenti in quantità inferiori o tracce sono ad esempio il stigmasterolo, 2%, colesterolo, brassicasterolo, ergosterolo.
Altre molecole presenti in quantità inferiori o tracce sono ad esempio il stigmasterolo, 2%, colesterolo, brassicasterolo, ergosterolo.
 Nella corsa il runner va incontro a un consumo calorico e a una perdita di acqua e sali minerali che dipendono da diversi fattori quali la tecnica, il grado di allenamento, le condizioni ambientali e le caratteristiche fisiologiche proprie di ciascun atleta. Ne consegue che la loro conoscenza permette di impostare un’alimentazione adeguata sia nella fase di allenamento che in quella di recupero tra un allenamento e il successivo, con l’obbiettivo di ottimizzare la performance.
Nella corsa il runner va incontro a un consumo calorico e a una perdita di acqua e sali minerali che dipendono da diversi fattori quali la tecnica, il grado di allenamento, le condizioni ambientali e le caratteristiche fisiologiche proprie di ciascun atleta. Ne consegue che la loro conoscenza permette di impostare un’alimentazione adeguata sia nella fase di allenamento che in quella di recupero tra un allenamento e il successivo, con l’obbiettivo di ottimizzare la performance.
 Il consumo limitato a questi quantitativi deve essere ottenuto con l’assunzione di bevande a basso contenuto alcolico, preferibilmente ai pasti (bere quantità anche piccole o moderate al di fuori dei pasti aumenta la probabilità di sviluppare ipertensione): ciò significa non più di 680 ml di una birra normale o 280 ml di vino (12% vol), soprattutto in caso di ipertensione; per le donne e i soggetti esili il consumo dovrebbe essere dimezzato1.
Il consumo limitato a questi quantitativi deve essere ottenuto con l’assunzione di bevande a basso contenuto alcolico, preferibilmente ai pasti (bere quantità anche piccole o moderate al di fuori dei pasti aumenta la probabilità di sviluppare ipertensione): ciò significa non più di 680 ml di una birra normale o 280 ml di vino (12% vol), soprattutto in caso di ipertensione; per le donne e i soggetti esili il consumo dovrebbe essere dimezzato1.
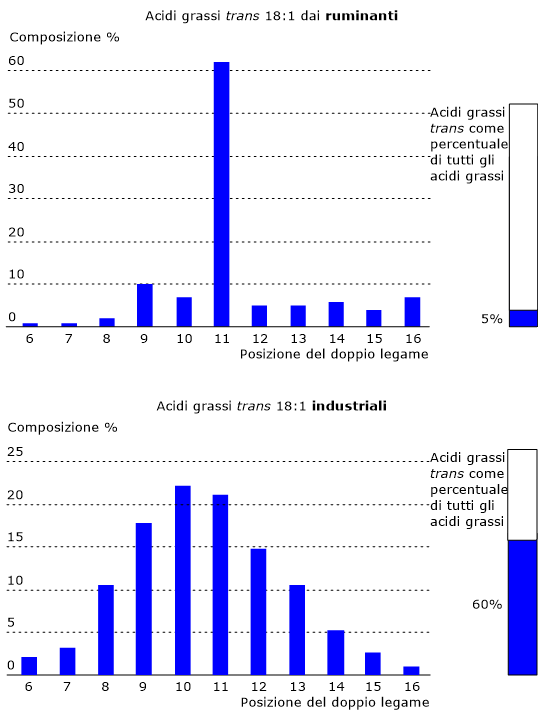
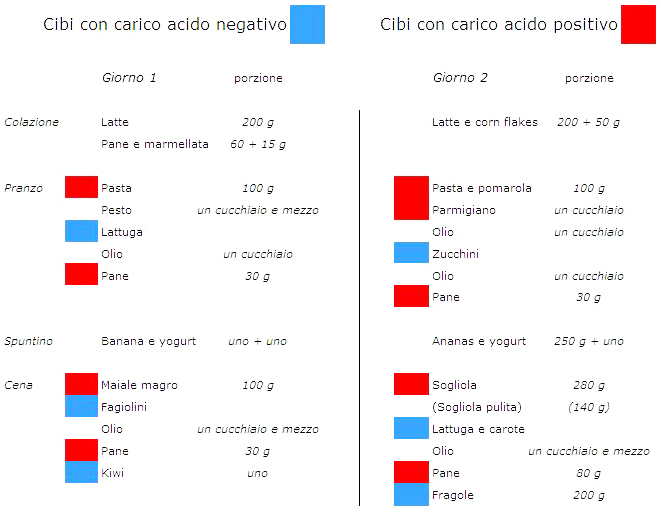

 Per produrre glicogeno è indispensabile assumere carboidrati; ma quanti, quali, quando e con che frequenza?
Per produrre glicogeno è indispensabile assumere carboidrati; ma quanti, quali, quando e con che frequenza?
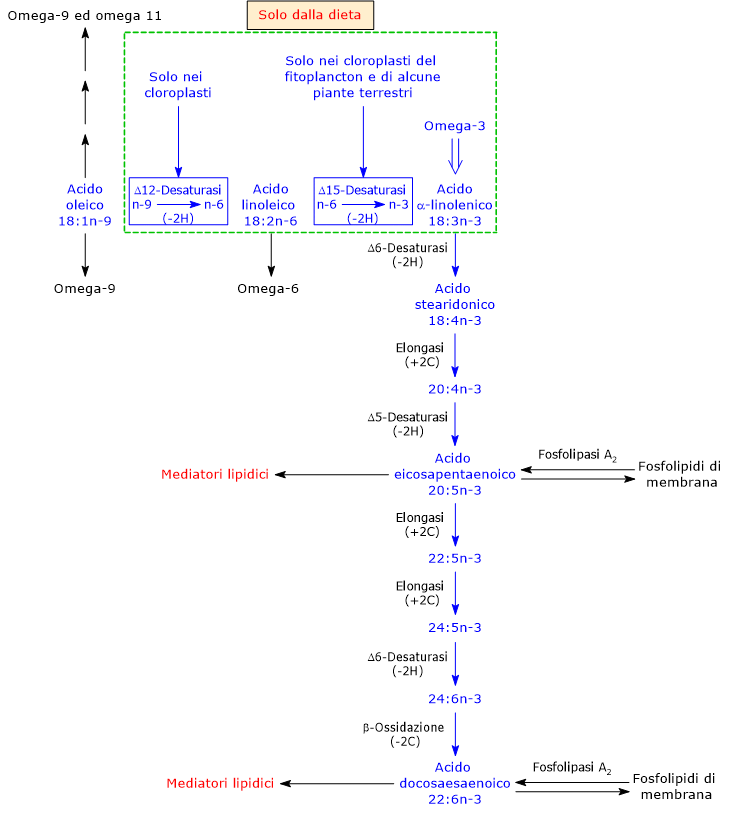
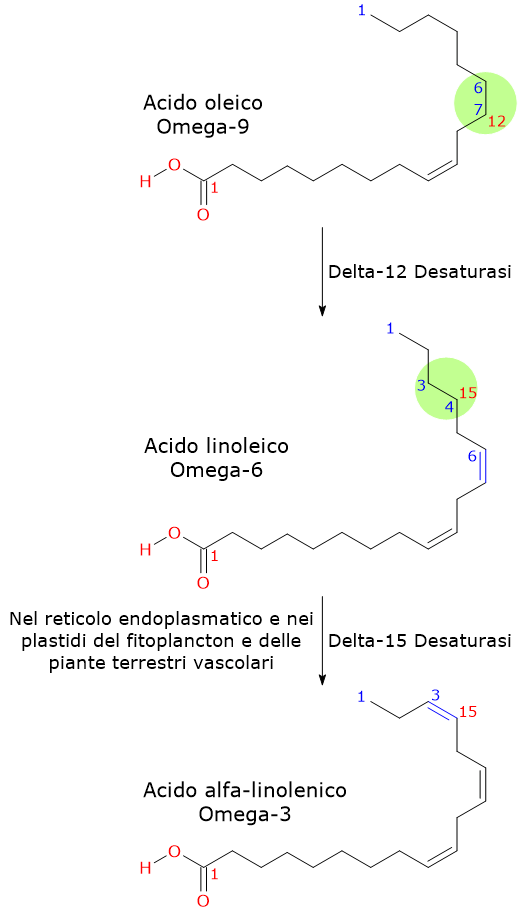
 Inoltre, a seconda del prodotto, può essere presente anche saccarosio non scisso.
Inoltre, a seconda del prodotto, può essere presente anche saccarosio non scisso.